
Hobbsinella gunnisonensis Camacho & Taylor, 2023
|
publication ID |
https://doi.org/10.5852/ejt.2023.891.2273 |
|
publication LSID |
lsid:zoobank.org:pub:BAB17D92-2A57-45FF-91D5-E3A3F3F6F10F |
|
DOI |
https://doi.org/10.5281/zenodo.8380351 |
|
persistent identifier |
https://treatment.plazi.org/id/A7F7AD89-20F6-49C3-91B9-4A89AB8BD83F |
|
taxon LSID |
lsid:zoobank.org:act:A7F7AD89-20F6-49C3-91B9-4A89AB8BD83F |
|
treatment provided by |
Plazi |
|
scientific name |
Hobbsinella gunnisonensis Camacho & Taylor |
| status |
sp. nov. |
Hobbsinella gunnisonensis Camacho & Taylor sp. nov.
urn:lsid:zoobank.org:act:
Figs 2–4 View Fig View Fig View Fig
Etymology
The species name, gunnisonensis (adjective, patronym), is derived from the Gunnison River Drainage Basin in the headwaters of the Colorado River where the new species occurs in two tributaries of the Gunnison River.
Material examined
Holotype USA • ♀; Colorado, Gunnison County, Lottis Creek off Forest Road 742; 38.77145° N, 106.62195° W; 3 Jul. 2018; C.B. Bonwell and J.N. McDonald leg.; dissection of all appendages and body parts preserved as permanent slides (special metal slides), glycerine-gelatine stained with methylene blue and paraffin as mounting medium; sample CO-137-4, CO-137-4-5; MNCN / ARTP20.04 /20172. GoogleMaps
Paratypes
USA • 5 ♀♀; same collection data as for holotype; dissection of all appendages and body parts preserved as permanent slides (special metal slides), glycerine-gelatine stained with methylene blue and paraffin as mounting medium; sample CO-137-4, CO-137-4-5; MNCN / ARTP20.04 /20173, MNCN / ARTP20.04 /20174, MNCN / ARTP20.04 /20180 ( MNCN / ADN54887 About MNCN ), MNCN / ARTP20.04 /20181 ( MNCN / ADN54888 About MNCN ), MNCN / ADN54889 About MNCN GoogleMaps • 1 ♀; same collection data as for holotype; dissection of all appendages and body parts preserved as permanent slides (special metal slides), glycerine-gelatine stained with methylene blue and paraffin as mounting medium; sample CO-137-3, CO-137-3-6; MNCN / ARTP20.04 /20179 ( MNCN / ADN54886 About MNCN ) GoogleMaps • 1 ♀; same collection data as for holotype; dissection of all appendages and body parts preserved as permanent slides (special metal slides), glycerine-gelatine stained with methylene blue and paraffin as mounting medium; sample CO-137-2, CO-137-2-4; MNCN / ARTP20.04 /20178 ( MNCN / ADN54885 About MNCN ) GoogleMaps .
Other material examined
USA • 3 ♀♀; Colorado, Gunnison County, Spring Creek off Forest Road 744; 38.75418°N, 106.76984° W; 3 Jul. 2018; C.B. Bonwell and J.N. McDonald leg.; dissection of all appendages and body parts preserved as permanent slides (special metal slides), glycerine-gelatine stained with methylene blue and paraffin as mounting medium; sample CO-136-3, CO-136-3-10; MNCN / ARTP20.04 /20175 ( MNCN / ADN54883 About MNCN ), MNCN / ARTP20.04 /20176 ( MNCN / ADN54884 About MNCN ), MNCN / ARTP20.04 /20177 GoogleMaps .
Description
MEASUREMENTS AND APPEARANCE. Body total length of holotype 1.12 mm. Total length of females 1.0– 1.21 mm. Body elongated, articles widening slightly towards posterior end, approximately ten times as long as wide. Head longer than wide. Pleotelson with one small barbed dorsal seta on each side.
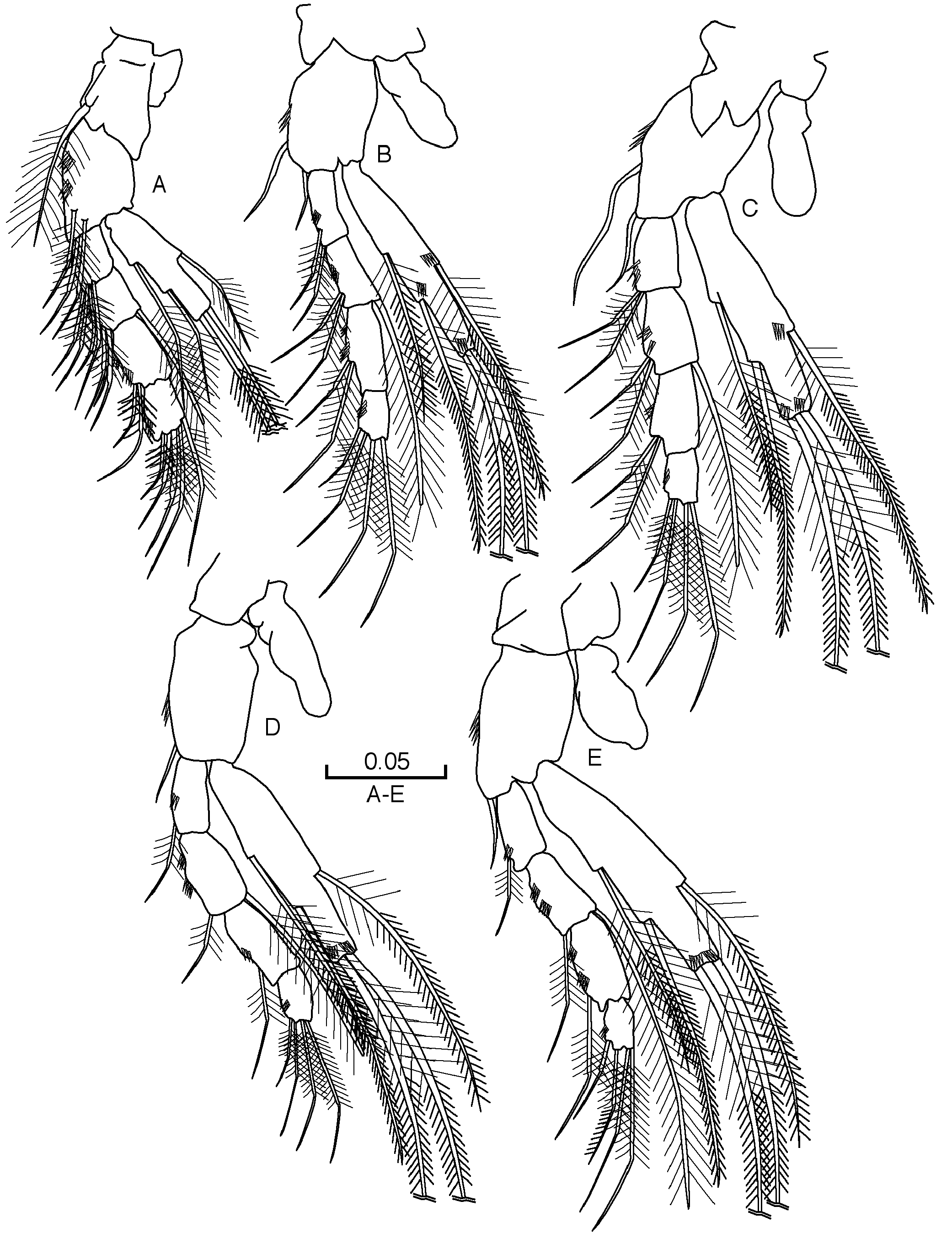
ANTENNULES (AI) ( Fig. 2A View Fig ). Seven-segmented; first three articles almost as long as last four articles combined; first article bit longer than the last article, which is more slender than the other articles; fourth article very short, fifth and sixth equal in length; inner flagellum almost square; setation as in Fig. 2A View Fig article six with three aesthetascs, similar in size; seventh article with three aesthetascs similar in size. AI slightly shorter than AII.
ANTENNAE (AII) ( Fig. 2B View Fig ). Seven-segmented; first article similar in length to the fourth; second and third articles are the shortest, while the fifth is slightly longer and just over the half length of the distal article setal formula: 0+0/1+0/2+1/2+0/0+0/2+2/5.
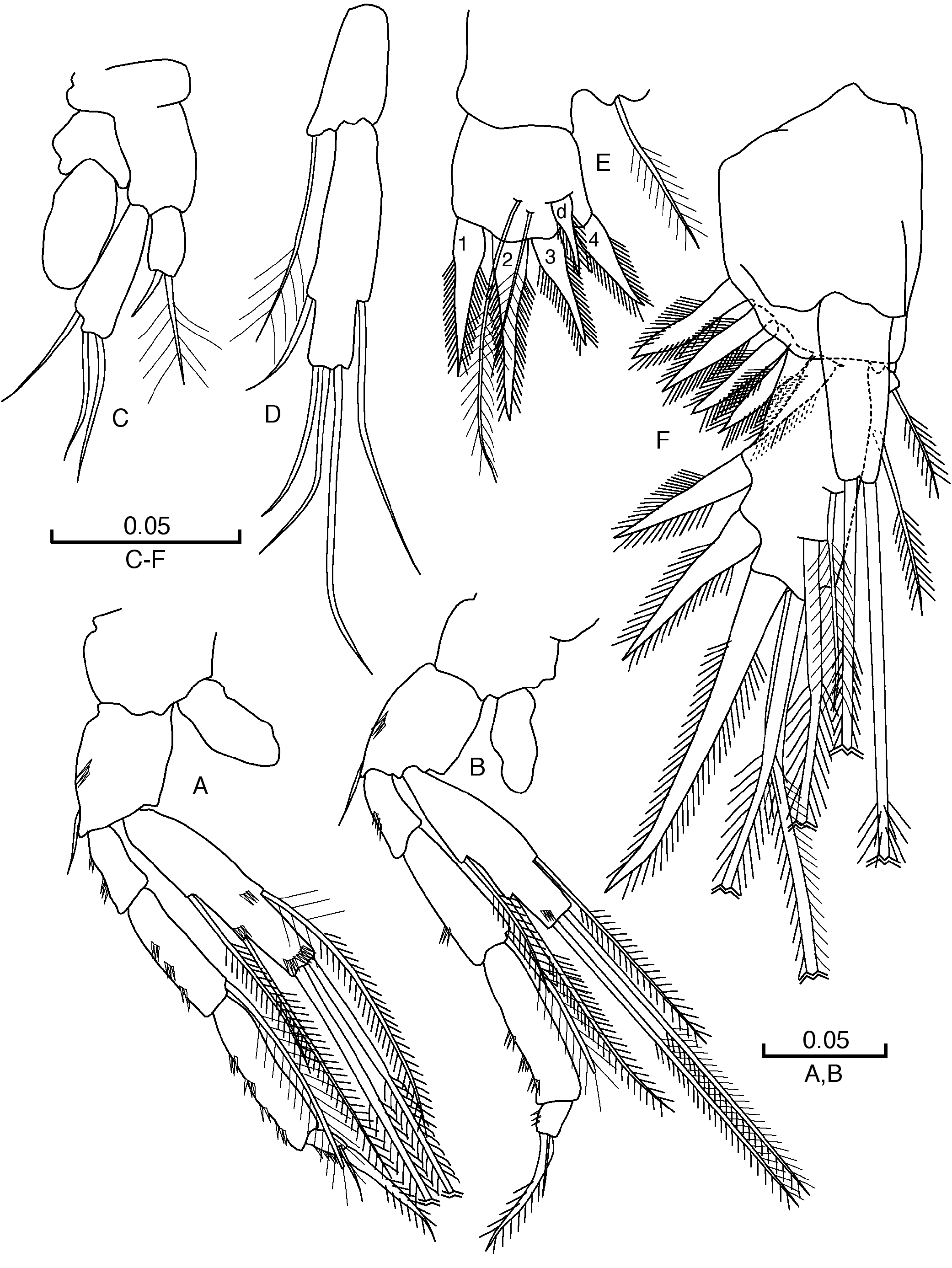
LABRUM ( Fig. 2C View Fig ). Almost trapezoidal; with smooth free edge and a median cleft.
PARAGNATHS ( Fig. 2D View Fig ). Almost rectangular, globose, with a very strong claw on distal part; dense setulation on distal half.
MANDIBLES (Md) ( Fig. 2E–F View Fig ). Palp with three articles, third article ( Fig. 2E View Fig ) with two strong barbed claws, first and third article almost square, second article elongated. Masticatory part ( Fig. 2E View Fig ): incisor process (pars incisiva) with two teeth; processus incisivus accessorius with one tooth and one small seta-like tooth; pars molaris with one tooth, nearest to processus incisivus accessorius, bidentate, and with two dentate structures, parallel to main axis of teeth, each with two small denticles and with a strong distal tooth.
MAXILLULES (MxI) ( Fig. 2G View Fig ). Proximal endite with four setae, three of them setulose; distal endite with six teeth (four with denticles and two seta-like); three plumose setae of different lenght, one longer than the other two, in outer margin.
MAXILLAE (MxII) ( Fig. 2H View Fig ). Four articles; setal formula 8, 5, 6, 6.
THORACOPODS (ThI to VII) ( Figs 3A–E View Fig , 4A–B View Fig ). Well developed ThI to III ( Fig. 3 View Fig A-C) progressively longer; ThIV ( Fig. 3D View Fig ) and V ( Fig. 3E View Fig ) of similar length; ThVI and VII ( Fig. 4A–B View Fig ) similar and a little longer than rest. Th I without epipod, coxa with long strong plumose seta, basipod with three smooth setae and tuft of long fine setules near base. Epipod present on ThII-VII, more than half length of basipod in all Ths. Exopod with one article in all Ths, shorter than endopod, similar in length to first three endopodal articles combined in ThI–V, slightly longer than first two articles combined in ThVI and VII, with five barbed setae (two terminal, one dorsal and two ventral). Endopod four-segmented in all thoracopods, all articles large and subequal in length in ThI to V; endopod of ThIII to V similar and slightly longer than in ThI–II and V, second and third articles very long in ThVI and VII; setal formula of endopods (number of setae on basipod in brackets):
ThI: (3) 4+0/3+1/2+0/5
ThII–III: (2) 2+0/2+1/2+0/4
ThIV–V: (1) 1+0/1+1/1+0/4
ThVI–VII: (1) 0+0/0+1/0+0/2(1)
THORACOPOD VIII (ThVIII) ( Fig. 4C View Fig ). Coxa without small seta; large and thick epipod, as long as basipod; endopod one-segmented with two unequal apical setae one smooth and one plumose; exopod two times as long as endopod, with three smooth similar setae, one of these subterminal and two other terminals.
PLEOPODS ( Fig. 4D View Fig ). Two segmented; first article with very long plumose seta; second article with five setae of different length.
PLEOTELSON ( Fig. 4E View Fig ). With one long, plumose dorsal seta at each side near base of furca.
FURCAL RAMI ( Fig. 4E View Fig ). Almost square, bearing five short spines of similar size except second, which is slightly longer than rest and the dorsal, which is slightly shorter.
UROPODS ( Fig. 4F View Fig ). Sympod, almost square, as long as endopod, with five long, equal distal spines; endopod almost 20% longer than exopod, with three strong claws (basal two subequal in length, distal claw twice as long as the others), three very long distal barbed setae ( Fig. 4I View Fig ) and one plumose setae located dorsolaterally; exopod with four setae (two terminal and two medial).
Remarks
The new species shares the combination of morphological characters listed in the diagnosis with the type species of the genus Hobbsinella , described originally from Texas ( Fig. 1B View Fig ): seven-segmented AI and AII; pars molaris of mandible with two parts; endopod of all thoracopods four-segmented; female thoracopod VIII biramous and a very large epipod. The genus presents some peculiarities with respect to other North American and European genera, as already highlighted when describing the genus from Texas ( Camacho et al. 2018b). The new species maintains those peculiarities of the type species but it is worth clarifying some of them. For example the fourth and last articles of AII in the species of Hobbsinella are longer than in other species of the American and European genera. The arrangement of the teeth on the pars molaris of the mandible is also unique. The female ThVIII has a very long epipod that exceeds the length of the basipod, but it is not bulky as in other genera with long epipods. The pleopod consists of two articles, as in all species of the family, but in most species, the first article is generally short (less than half the length of the second). In Hobbsinella , the first article of the pleopod is two thirds the length of the second. The spines of the sympod of the uropod are fairly long and not very thick, unlike most genera with shorter, thicker sympodal spines. The morphological differences between the species are difficult to find. The new species is slightly smaller than the type species, H. edwardensis . Hobbsinella gunnisonensis sp. nov. AII is slightly longer than AI (as Vandelibathynella Serban, Coineau & Delamare Deboutteville, 1971 ) while in H. edwardensis the great length of the AII is very striking, which far exceeds the length of AI ( Table 3 View Table 3 ) ( Camacho et al. 2018b). The third article of AI has only three setae on the new species, five in H. edwardensis , and also three aesthetacs on article six (only two in the type species), but the setal formula of the rest of articles is similar in both species and both species lacks of medial seta on exopod of AII. Both species differs also in the combinations of setae on the articles of the endopod of ThI to V, as well as the combinations of setae on the basipod of all thoracopods ( Table 3 View Table 3 ). The differences between the two species are subtle and very difficult to appreciate, since in general they refer to the size, appearance and relative proportions of the different articles. The number of spines and/or setae on MxII, Ths and uropods differ between the two species, and these differences are summarized ( Table 3 View Table 3 ) to facilitate comparison.
Molecular results
18S rRNA and COI sequences were obtained from five females specimens of the new species ( Table 2 View Table 2 ).
The concatenated COI–18S data set is represented by 31 sequences of 1580 bp. 509 bp COI and 1071 bp 18S sequences were obtained from 31 specimens.
The uncorrected sequence divergence estimates for 18S between genera and the outgroup within the family Bathynellidae are consistent with previous studies ( Camacho et al. 2016, 2018 a, 2018 b, 2020, 2021; Perina et al. 2019a, 2019b). For 18S, sequence divergence between the genera of Bathynellidae and the outgroup used in the phylogeny ( Iberobathynella imuniensis Parabathynellidae family) ranged between 7.4–8.2% ( Gallobathynella ), 8.0% ( Altainella) and 12.3% ( Pilbaranella ). Hobbsinella shows a 18S divergence range with Gallobathynella and Vejdovskybathynella of 5.4%–5.7% and 5–6%–7.8% with Paradoxiclamousella and 9.0%–15.4% with Australian genera ( Perina et al. 2019a).
The uncorrected sequence divergence estimates for COI between the new species and H. edwardensis is 8.5–8.7% and is consistent with the values found between congeneric species such as Vejdovskybathynella edelweiss Camacho, 2007 (different populations), V. vasconica Camacho, Dorda & Rey, 2013 (14.7%) and V. caroloi Camacho, 2007 that shows a COI divergence ranged between 6.5–7.5% or between B. ruffoi Serban, 1973 and other European Bathynella undetermined (4.6–6.2%). Between populations of the new species the difference found is 0.3%, less than between populations of V. edelweiss (0.5– 0.6%).
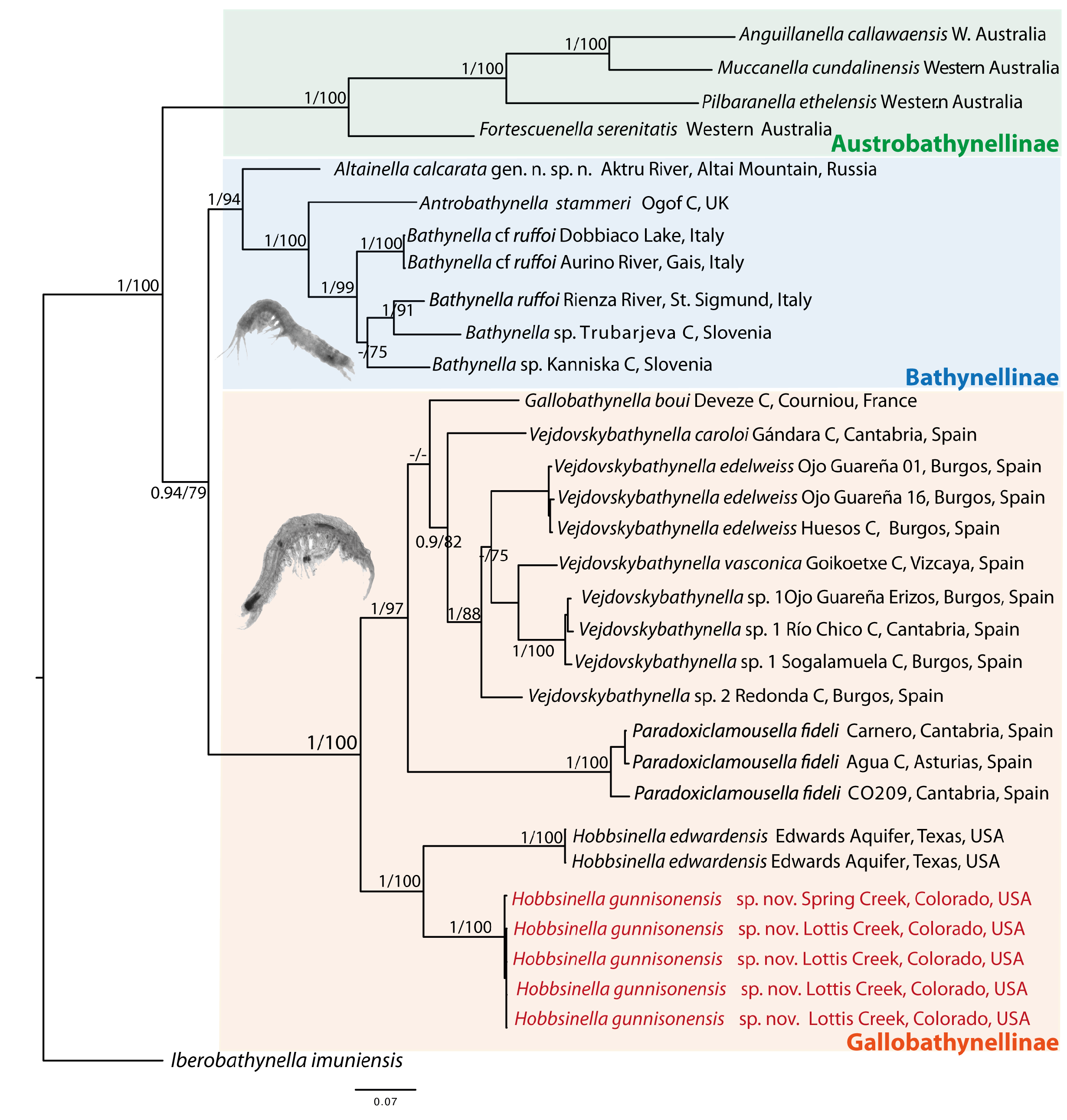
Maximum Likelihood (ML) (bootstrap support, BS) and Bayesian Inference (BI) (posterior probabilities, PP) phylogenetic analyses recovered similar topologies ( Fig. 5 View Fig ), supporting two monophyletic clades corresponding to genera of Bathynellidae from Australia and the other genera (PP = 0.94; BS = 79). European and North American genera form two well supported, monophyletic lineages (PP =1;BS = 100). One of these correspond to the subfamily Bathynellinae Grobben, 1905 and the other to the subfamily Gallobathynellinae Serban, Coineau & Delamare Deboutteville, 1971 in which the new species is placed. The subfamily Gallobathynellinae comprises species of the genera Vejdovskybathynella (PP = 0.9, BS = 82) and Paradoxiclamousella (PP = 1, BS = 100) from the Iberian Peninsula, Gallobathynella from France and Hobbsinella (PP = 1, BS = 100) from the USA with two well differentiated and supported clades (PP = 1, BS = 100). One clade includes H. edwardensis from Texas and the other includes the specimens of H. gunnisonensis sp. nov. from the two localities in Colorado. The subfamily Bathynellinae includes species of Bathynella from Slovenia and Italy; Antrobathynella stammeri (Jakobi, 1954) from the UK and Altainella calcarata Camacho et al., 2020 from Russia.
The four species described from Western Australia ( Anguillanella callawaensis Perina & Camacho, 2019 , Fortescuenella serenitatis Perina & Camacho, 2019a , Muccanella cundelinensis Perina & Camacho, 2019 and Pilbaranella ethelensis Perina & Camacho, 2018 ) could represent Austrobathynellinae Delamare Deboutteville & Serban, 1973, but molecular data of the original species for which the subfamily was created are needed to confirm this.
Molecular data support morphology and the decisión to create a new species


| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Gallobathynellinae |
|
Genus |