
Pseudosinella gisini Christiansen, 1961
|
publication ID |
https://doi.org/10.5281/zenodo.275457 |
|
persistent identifier |
https://treatment.plazi.org/id/0B21878A-6374-FD18-F7EF-CFC0FD0788EB |
|
treatment provided by |
ImsDioSync (2017-06-17 15:55:21) |
|
scientific name |
Pseudosinella gisini Christiansen, 1961 |
| status |
|
Pseudosinella gisini Christiansen, 1961
Figs 71–86 View FIGURES 71 – 80 View FIGURES 81 – 84 View FIGURES 85 – 86
Material Examined. GREENBRIER CO.: Al’s Insurgence, 28 May 2004, 1 alcohol; Allison's Cave, 29 May 2004, 1 slide, 1 alcohol; Benedicts Cave System Persinger’s Entrance, 1.June. 2004, 36 in alcohol; Bill Jones, DATE, 2 alcohol; Boar Hole, 18 June 2004, 7 alcohol; Boothe Cave, 12 June 2004, 1 alcohol; Bubble Cave, 5 May 2004, 18 alcohol; Upper Buckeye Creek Cave, 18 June 2004, 30 alcohol; Callison’s Pond, 22 May 2004, 5 alcohol; Clutetown Cave, 22–25 June 2004, 16 alcohol; Court Street Cave, 28 May 2004, 2 alcohol; Culverson Creek System, Hinkle-Unus Entrance, 1 in alcohol; Destitude Cave, 4 in alcohol; Field Station Pit, 6 June 2004, 3 alcohol; Fox Cave, 30 May 2004, 6 alcohol; Fuells Fruit Cave, 2 on slides, 17 in alcohol; General Davis Cave, 6 in alcohol; Grape Vine/Lost World Caverns, 28 May 2004, 1 slide, 19 alcohol; Goat Cave, 11 June 2004, 1 alcohol; Hell of a Pit 2, 28 May 2004, 1 on slide, 16 in alcohol; Hillside Pit, 9 June 2004, 12 in alcohol; Hit N Head Cave, 25 June 2004, 1 alcohol; The Hole System Gibbs Entrance, 6 alcohol; Inspired Pit, 5–8 June 2004, 1 alcohol; Ludington's Cave, 15 June 2004, 4 alcohol; MC Cave, 1 June 2004, 1 slide, 3 alcohol; MC Pit, 2 June 2004, 6 alcohol; McFerrin Water (Spur), 1 alcohol; McFerrin Breakdown Cave, 22 August 2004, 7 alcohol; Nellie’s Cave, 20–23 May 2004, 4 alcohol; Osborne Pit, 2 June 2004, 3 on slides, 40 in alcohol; Organ Cave System, Lipp Entrance, 21 May 2004, 1 slide, 4 alcohol; Oak Sang Cave, 14 June 2004, 3 alcohol; Pilgrims Rest Church, 19 alcohol; Pollock Cave, 1 on slide, 1 head and 1 leg of a second specimen in alcohol; Raceway Pit, 31 May 2004, 8 alcohol; Rapps Cave, 28 May 2008, 2 alcohol; Salamander Suicide Pit, 2 June 2004, 18 alcohol; Seep Cave, 12 June 2004, 4 alcohol; Spencer Cave, 17 June 2004, 1 alcohol; Upper Spout Cave, 3 alcohol; Teetering Rock Cave, 16 June 2004, 6 alcohol; Tin Cave, 9 June 2004, 1 alcohol; Trillium Cave, 28 May 2004, 1 alcohol; Turner Pit 2, 1 on slide; US 219 Cave, 27 May 2004, 10 alcohol; Zimmerman's Pit, 11 June 2004, 7 alcohol; MONROE Co: Greenville Saltpeter Cave, Mill Pond Entrance, 10 June 2004, 2 alcohol; Hurricane Ridge Cave, 6 June 2004; Steele’s Cave, 11 June 2004, 1 on slide; Union Cave, 1 June 2004, 2 alcohol; POCAHONTAS Co: Blue Spring Cave, 10 June 2004, 2 slides, 74 alcohol; Lobellia Saltpeter Cave, 6 June 2004, 29 alcohol; Stella’s Cave, 14 June 2004, 11 alcohol; Martha’s Cave, 20 August 2004, 2 slides, 7 alcohol.
Descriptive notes. Color in alcohol variable among different caves and instars, from white, completely unpigmented, to light blue pigment only on eye patch or light blue in the form of small dots uniformly distributed through out body. Manubrial length of the individuals examined ranging from 0.26–0.80mm. Individuals with manubrium measuring 0.71mm or less show a range of variable conditions probably attributable to instability of characters during postembryonic development ( Szeptycki 1979, Soto-Adames 2008).
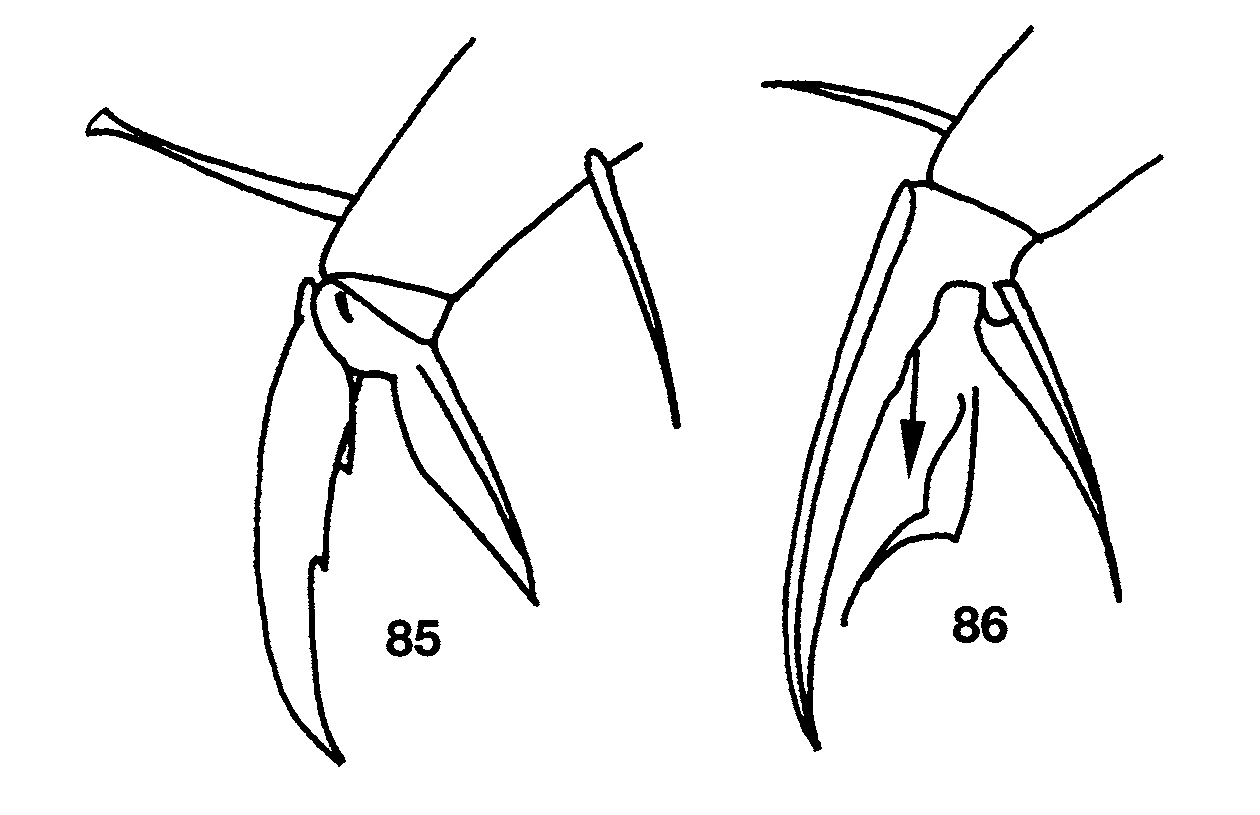
Ant. 4 subapical sense organ capitate ( Fig. 71 View FIGURES 71 – 80 ). Ant. 3 sense organ formed by 2 thin walled sensilla varying from somewhat expanded laterally to paddle-shaped; accessory sensilla as in Fig. 74 View FIGURES 71 – 80 , sensilla 7 a present in the largest individual with Ant. 3 (manubrium=0.70 mm long). Macrosetae at base of antennae variyng from 5–11, larger individuals usually with more macrosetae but correlation between manubrial size and setae number not perfect. Dorsal head chaetotaxy ( Fig. 72 View FIGURES 71 – 80 ) with macrosetae A0, A 2, and A 3; setae A 2 a enlarged, shorter than, but with socket not different in size from macroseta A 2 ( Fig. 73 View FIGURES 71 – 80 ); Pa 5 a microseta. All dorsal microsetae ciliate: M0 present; M 1 /S 1 closer to M 2 than S0; row S with 5 + 1 setae, S0 anterior to S 3. Eyes 2 + 2, subequal. Prelabral and labral setae smooth. Inner labral papillae smaller than outer papillae, all papillae smooth. Pleural fold setae smooth. Peristomal seta pss 0 smooth or ciliate, pss 1–2 smooth and sharply acuminate. Maxilla with 4 ungular teeth ( Fig. 80 View FIGURES 71 – 80 ). Labial papilla E with lateral process curved dorsally and just reaching tip of papilla. Basal proximal seta of labial palpi with seta Y subequal or slightly longer than Z. Chaetotaxy of labial triangle with M 2 always smooth, but otherwise variable, with five combinations of characters involving variation in sculpturing of M 1, r, E and L 1 depending on size of individual ( Figs 78–79 View FIGURES 71 – 80 ). All individuals with manubrium ≥ 0.71 mm have all posterior labial setae smooth (i.e., M 1 M 2 rEL 1 L 2). At smaller manubrial sizes variation pattern not clear: all individuals with manubrial length ≥ 0.60mm have seta M 1 smooth and all individuals with manubrium ≤ 0.29mm have M 1 ciliate, but M 1 condition not predictable in individuals between these size ranges. Likewise, r is smooth at manubrial sizes ≥ 0.71mm, ciliate at sizes ≤ 0.29mm and unpredictable at intermediate sizes. Labial setae L 1 and E ciliate only in one small individual (manubrium = 0.47mm) in which M 1 and r are also ciliate. Postlabium ( Fig. 78 View FIGURES 71 – 80 ) with 4 + 4 setae along ventral groove, seta 1–3 smooth, seta 4 ciliate; columns M, E, L and O with 2, 2, 3 and 2 setae respectively, all setae on anterior row (except O 1) and O 2 smooth, setae C 2 and L 3 ciliate, E 2 smooth or ciliate, setae L 2 and O 1 ciliate and shorter than other postlabial setae. Formula for inner macrochaetotaxy of body as 00/0100+ 2. Mesothorax moderately enlarged. Abd. 1 setae organization linear; seta a 6 present. Chaetotaxy of Abd. 2 ( Fig. 81 View FIGURES 81 – 84 ) with all supplementary setae fan-shaped; all other setae (except as) ciliate, a 2 and a 3 sometimes distally expanded but not quite fan-shaped (inset Fig. 81 View FIGURES 81 – 84 ); 2–3 setae in series m around inner bothriotricha, a 2 p present; a 3 inserted anterior to as, usually not reaching as, but long and surpassing as in some individuals ( Fig. 81 View FIGURES 81 – 84 inset); as may or may not reach socket of m 3; m 3 e not reaching socket of m 3; series L anterior to lateral bothriotricha with 2 setae; m 5 a normal macroseta; p 5 p present and longer than p 5. Abd. 3 ( Fig. 82 View FIGURES 81 – 84 ) with a 2, a 6, am 6 and all supplementary setae fan-shaped, all other setae (except as and d 2) ciliate; in most individuals a 3 inserted between as and a 2 and not reaching as, in others, a 3 inserted between m 3 and as and reaching base of as ( Fig. 82 View FIGURES 81 – 84 inset); as shorter than, but reaching m 3; d 2 present; a 7 ciliate and lanceolate and inserted very close to am 6; m 7 and p 7 normal microsetae, m 7 inserted between pm 6 and p 6, m 7 a an acuminate macroseta. Abd. 4 bothriotrichal complex ( Fig. 83 View FIGURES 81 – 84 ) with m, a, s, and D 1 fan-shaped; s smaller than surrounding supplementary setae; C 1 p, T 3 and D 1 p ciliate; tip of T 3 reaching base of D 1 p; D 1 p posterior to T 3 and almost reaching Pe; Pe and Pi fan-shaped. General chaetotaxy of Abd. 4 ( Fig. 84 View FIGURES 81 – 84 ) with macrosetae B 5, B 6, T 6, T 7, D 2, D 3, E 1, E 2, E 3, F 1, F 2 and F 3; macroseta B 5 crossed by, or just anterior to a line drawn between A 5 and C 2; a supplementary microseta present between B 5 and B 6 seen in 14 of 21 individuals examined (arrow in Fig. 84 View FIGURES 81 – 84 ), presence of seta not correlated with manubrial size; F 2 closer to E 2 than E 3; microseta posterior to E 3 present. Abd. 4 with 5 + 5 posterior setae. Macrosetae of metathoracic femora not seen although sockets clearly enlarged. Claw complex showing two distinct morphologies. Most individuals with tenet hair spatulate ( Fig. 85 View FIGURES 85 – 86 ), subequal to slightly shorter than unguiculus; unguiculus weakly truncate to lanceolate, with basal swelling varying in development; posterior lamella serrate in some individuals; unguis with 3 inner and 3 outer teeth, inner basal pair unequal in size, outer tooth very short on prothoracic legs, but distinct on metathoracic legs; lateral teeth short on prothoracic legs but apparently absent from metathoracic legs. One individual from Fuell’s Fruit Cave and one from Scott’s Hollow Cave have an elongate unguis ( Fig. 86 View FIGURES 85 – 86 ) with only the basal pair of inner teeth present, the outer tooth is very close to the base of the unguis and the lateral teeth appear to be absent, the tenent hair is acuminate and shorter than the unguiculus, and the unguiculus is sharply acuminate. These two individuals are relatively small (manubrial length≈ 0.47mm for individual from Fuell’s Fruit Cave), either have one or no eyes (Fuell’s Fruit Cave), and have all peristomal and setae (except pss 2) ciliate ( Fig. 75 View FIGURES 71 – 80 ), but otherwise are similar to other individuals in the samples in which they were included. Ventral tube of most individuals with 2–3 + 2–3 macrosetae on distal anterior margin and 16 disto-lateral setae, three small individuals (manubrium ≤ 0.29mm) with only one distal macroseta on anterior face; setae on posterior face obscured in all specimens studied, but apparently more than ten in number. Chaetotaxy of manubrial plate with 6 outer and 2 inner setae separated by 2 pseudopores. Apical mucronal tooth not much longer than basal tooth.
Remarks. Christiansen and Bellinger (1996) divided P. g i s i n i into three subspecies based on the presence or absence of microsensilla 7 a on the sense organ of Ant. 3, eye number, sculpturing of setae along the ventral groove on the head (postlabial setae I), and to a lesser extent the labial and ventral tube chaetotaxy. Thus, the presence of sensilla 7 a on the sense organ of Ant. 3 identifies P. g. gisini , absence of sensilla 7 a and reduction of labial seta r identifies P. g. virginia Christiansen & Bellinger, 1996 and the combination of an absent sensilla 7 a and ciliate labial seta r characterizes P. g. carolina Christiansen & Bellinger, 1996. Other characters show uneven degrees of overlap among the three forms and can be used to differentiate among them only in combination with the characters mentioned above. Unfortunately, in 10 of the 21 individuals examined Ant. 3 had broke off or was obstructed by other appendages, but the individuals in which the presence or absence of sensilla 7 a could be determined form an heterogeneous group and do not fit the range of variation proposed by Christiansen and Bellinger (1998) for each subspecies. Of the 11 individuals in which Ant. 3 is intact, 5 have sensilla 7 a and would be identified as P. g. gisini , whereas 6 individuals have a small acuminate labial seta r which is either ciliate or smooth and would be identified as P. g. c a ro l i n a. However, all putative P. g. carolina have 2 eyes (except the individual from Fuell’s Fruit Cave mentioned above in reference to claw structure, which is blind) instead of the 0–1 typical of the subspecies, and all have the anterior seta in postlabial column I smooth (ciliate in typical P. g. c a ro l i n a). Following the geographical distribution of the subspecies reported by Christiansen & Bellinger (1998), all individuals examined here should belong into P. g. gisni and all should have sensilla 7 a. Most individuals in which only one 7 -series sensilla is visible are small and it is possible that either 7 or 7 a is secondary and absent in early instars (cf. Figs. 74, 76–77 View FIGURES 71 – 80 ). The absence of sensilla 7 a in medium size individuals may result from broken antennae that did not re-grow a normal complement of sensilla.
Christiansen, K. (1961) The genus Pseudosinella in caves of the United States. Psyche, 67 (1960), 1 - 25.
Christiansen K. & Bellinger P. (1996) Cave Pseudosinella and Oncopodura new to science. Journal of Caves and Karst Studies, 58, 38 - 53.
Christiansen K. & Bellinger P. (1998) The Collembola of North America north of the Rio Grande; A taxonomic analysis. Grinnell College, Grinnell, Iowa, 1518 pp.
Szeptycki, A. (1979) Chaetotaxy of the Entomobryidae and its phylogenetical significance. Morpho-systematic studies of Collembola, IV. Polska Akademia Nauk, Zaklad Zoologii Systematycznej i Doswiadczalnej, Krakow, Poland, 219 pp.
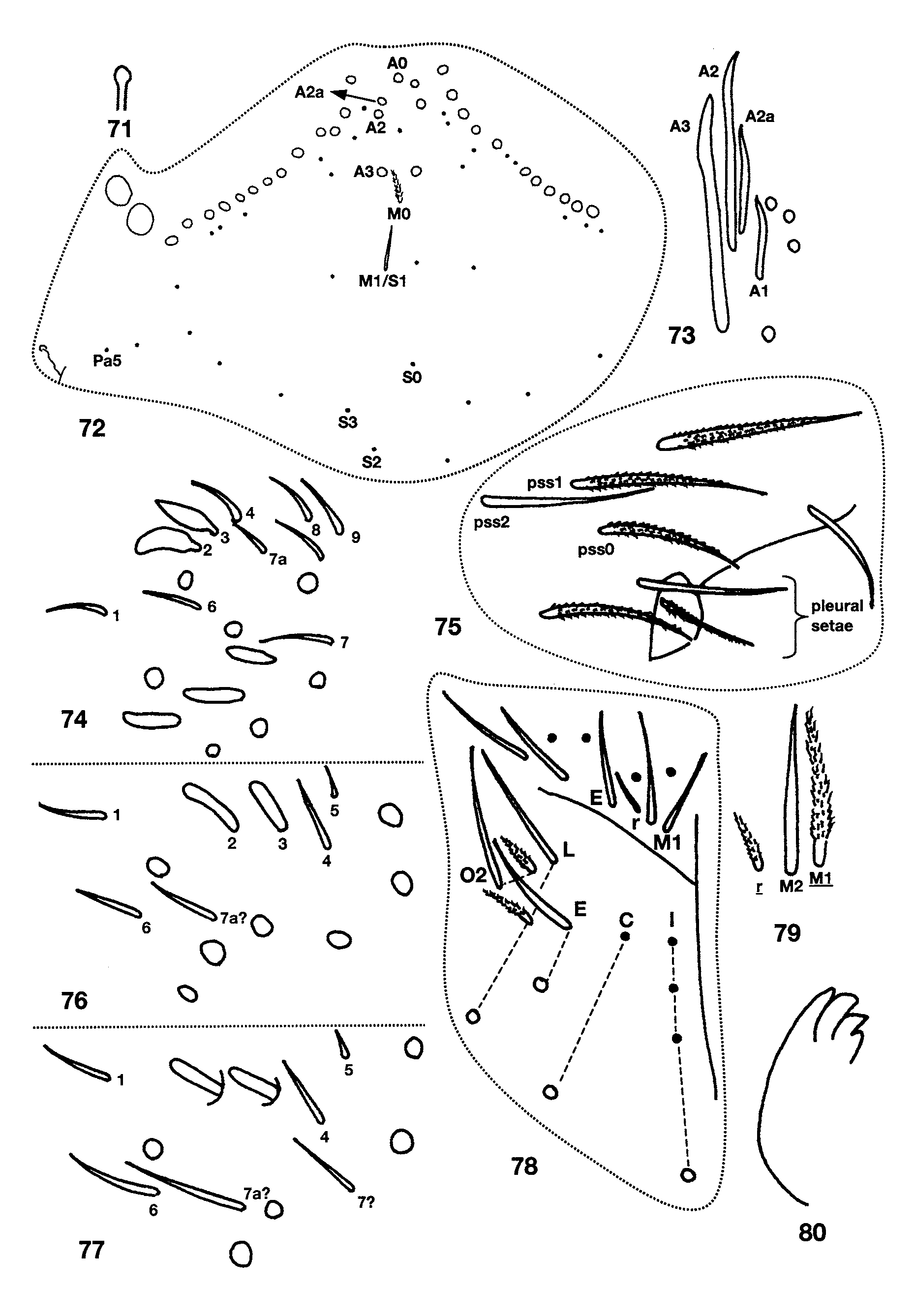
FIGURES 71 – 80. Pseudosinella gisini: 71, Subapical sensilla of fourth antennal segment; 72, Dorsal head chaetotaxy; 73, Detail of dorsal head macrosetae on series A showing development of seta A 2 a and relative size of socket; 74, Normal sense organ of third antennomere and associated sensilla, view of sensillum 5 obstructed by sensillum 3; 75, Peristomal and pleural setae in small juvenile; 76, Sense organ of third antennomere and associated sensilla in small juvenile; 77, Atypical sense organ of third antennomere and associated sensilla; 78, Labial and postlabial chaetotaxy; 79, Alternative morphology of labial setae m and r; 80, Ungulum of maxilla.
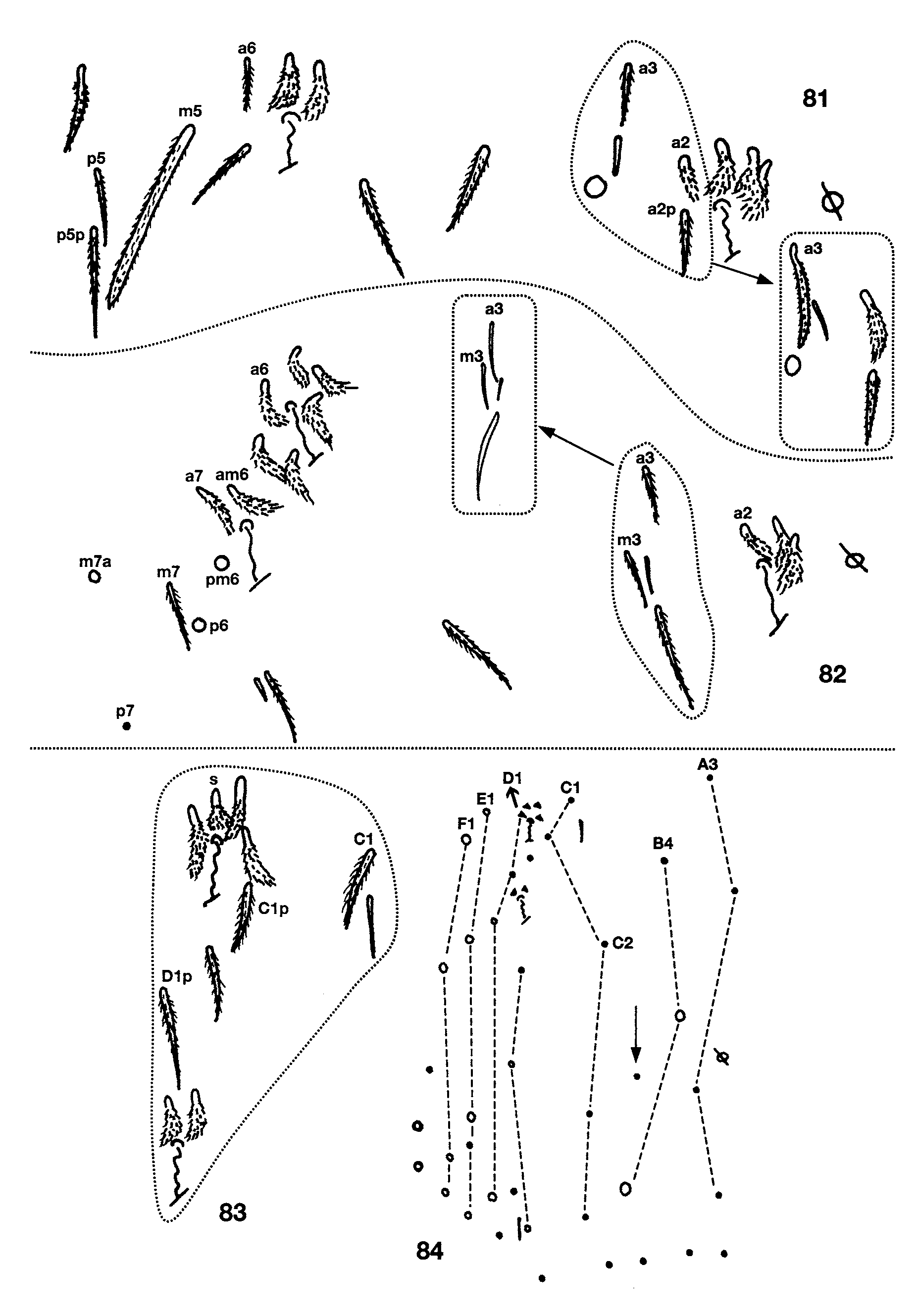
FIGURES 81 – 84. Pseudosinella gisini: Abdominal chaetotaxy, circles, dots and triangles are macro-, micro and fanshaped setae, respectively. 81, Second segment; 82, Third segment; 83, Bothriotrichal complex of fourth segment; 84, Complete chaetotaxy of fourth segment.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
1 (by plazi, 2016-04-08 05:23:07)
2 (by ImsDioSync, 2017-06-17 15:55:21)
3 (by ImsDioSync, 2017-06-17 20:09:27)
4 (by ExternalLinkService, 2019-09-26 07:03:05)
5 (by ExternalLinkService, 2022-01-30 17:01:26)
6 (by ExternalLinkService, 2022-02-21 18:04:21)
7 (by plazi, 2023-10-25 11:52:45)