
Travassosinema claudiae, Morffe & Hasegawa, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4282.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:D853A61B-EE41-44C5-9C62-C221081326D8 |
|
DOI |
https://doi.org/10.5281/zenodo.6049679 |
|
persistent identifier |
https://treatment.plazi.org/id/0B4187CB-1026-8259-FF58-F9601759FAAE |
|
treatment provided by |
Plazi |
|
scientific name |
Travassosinema claudiae |
| status |
sp. nov. |
Travassosinema claudiae n. sp.
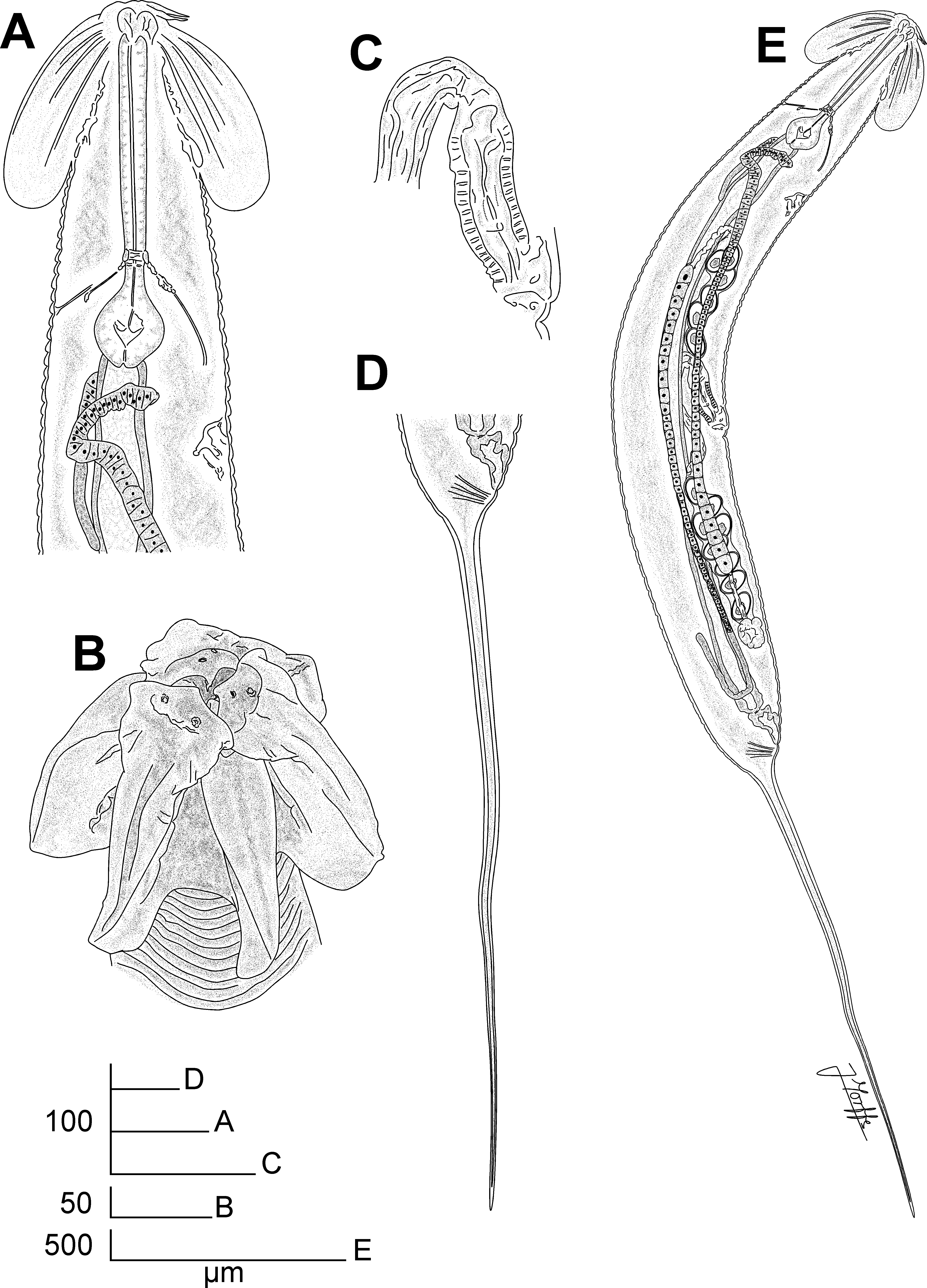
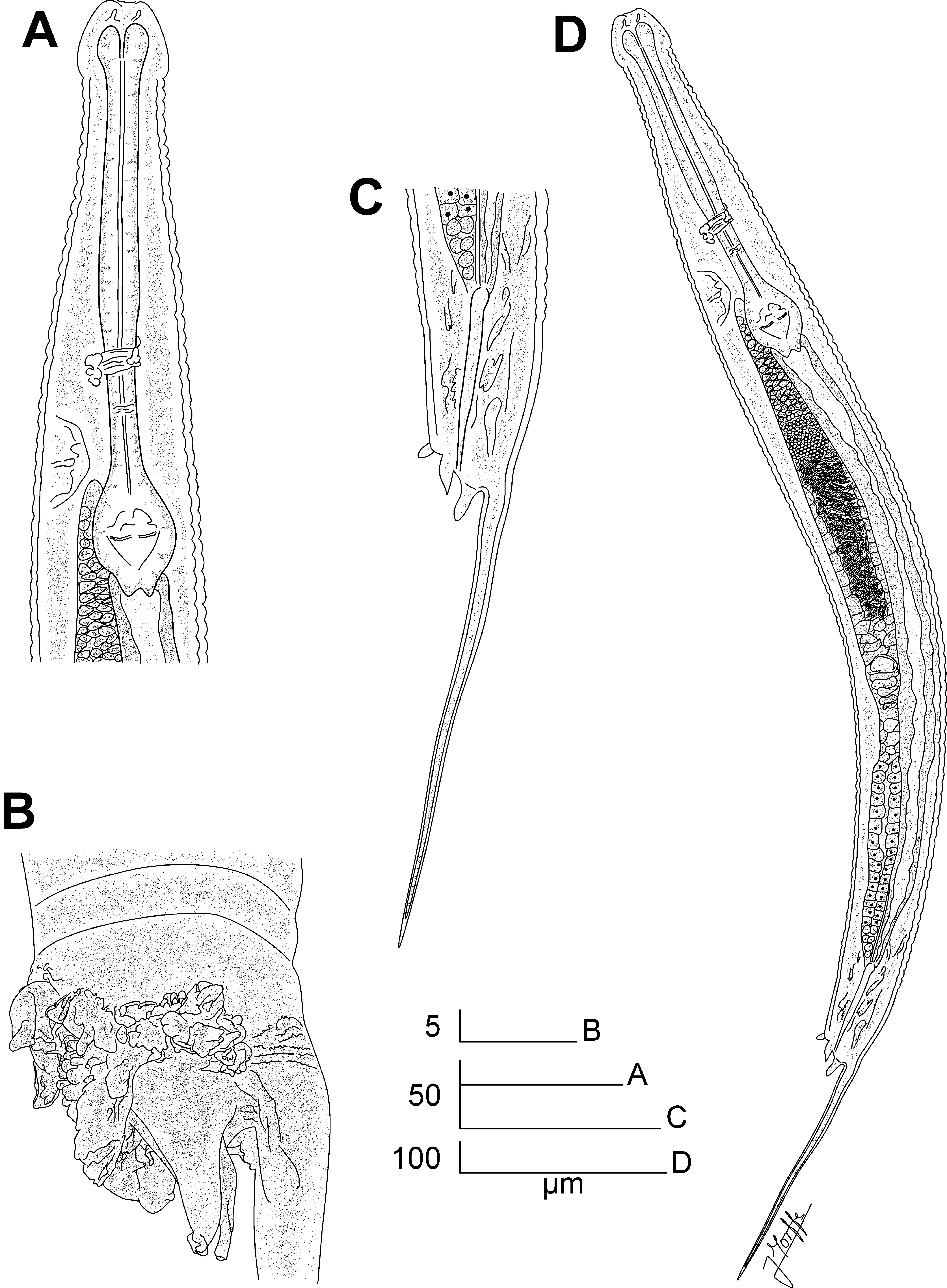
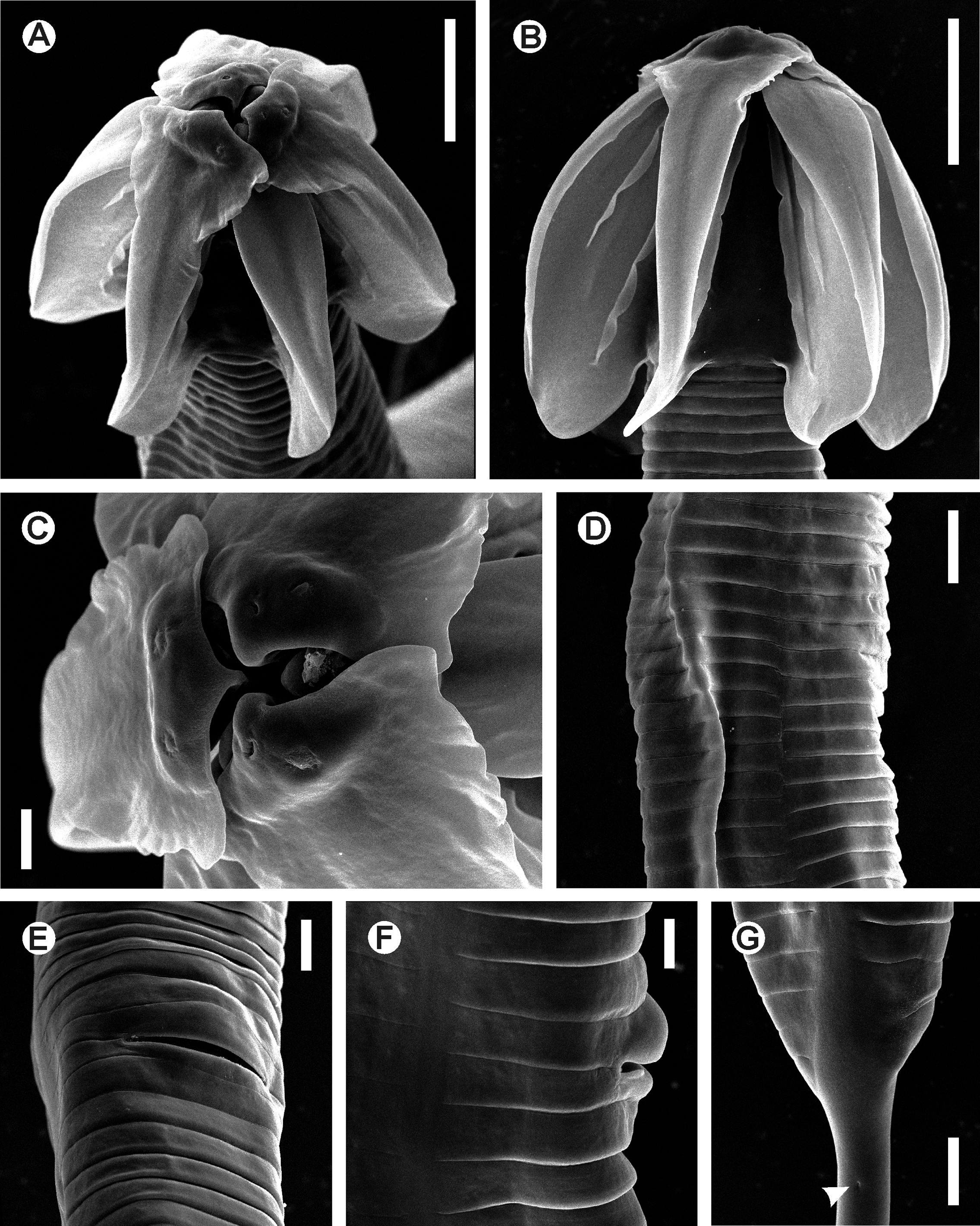
Fig. 1 View FIGURE 1 A–E, Fig. 2 View FIGURE 2 A–D, Fig. 3 View FIGURE 3 A–G, Fig. 4 View FIGURE 4 A–F
Type material. Holotype: ♀, Japan, Aichi prefecture, Kasugai, Chubu University Campus ; in Parafontaria laminata ; 20/VI/2016; S. Ozawa, K. Sato, J. Morffe coll.; CZACC 11.7016 View Materials . Paratypes: 7♀♀, Japan, Aichi prefecture, Kasugai, Chubu University Campus ; in Parafontaria laminata ; VII/2016; J. Morffe coll. ; CZACC 11.7017–11.7023 . 5♂♂, same data as the latter ; CZACC 11.7024–11.7028 . 1♀, Japan, Aichi prefecture, Kasugai, Chubu University Campus ; in Parafontaria laminata ; 20/VI/2016; S. Ozawa, K. Sato, J. Morffe coll. ; NCHL-A 13 . 1♀, Japan, Aichi prefecture, Kasugai, Chubu University Campus ; in Parafontaria laminata ; VII/2016; J. Morffe coll. ; NCHL-A 14 . 2♂♂, same data as the latter ; NCHL-A15, A16.
Measurements. See Table 1.
Description. Female. Body robust, spindle-shaped, with conspicuous umbraculum in the cephalic region. Cuticle markedly annulate from posterior end of the umbraculum to level of the anus. A number of ca. 54 annuli present before the level of the vulva, its width ca. 5 µm near the cephalic umbraculum, then increasing their diameter to ca. 10 µm towards the level of the vulva. Annuli posterior to the vulva in number of ca. 28, its width ca. 10 µm. Lateral alae absent. Lateral fields present, as a rupture of the annuli, starting from a short distance posterior to the oesophagus (ca. the 20th annuli posterior the umbraculum) to the level of the anus. Rupture of the annuli is evident in anterior part of the lateral field as a discontinuity with lateral displacement of the annuli, posteriorad along the body the rupture becomes wider. Oral opening triradiate, with three well developed, slightly overlapped lips, arranged as one dorsal and two sub-ventral. Three rounded interlabia located beneath the level of the lips, arranged as one ventral and two sub-dorsal. Four cephalic papillae present, two on the dorsal lip and one on each of the sub-ventral lips. Amphids lateral, C-shaped, located one on each of the sub-lateral lips, dorsal to the cephalic papillae. Cephalic umbraculum consisting of six radially arranged and posteriorly directed alae, arising from each labium and interlabium and with their rounded posterior ends extending to ca. two thirds of the procorpus length, with the point of attachment to the body ca. five annuli anterior. Edges of alae strongly convex, with the cuticle laterally extended, forming a hood-like structure, wider in the apical part of the alae and diminishing towards their posterior ends until they disappear near the level of the somatic attachment. This hoodlike structure is more developed in the apical end of the lip elements, being wider than the interlabial elements and with the margins sinuated. Each umbraculum ala is supported by three cuticular ribs of different length: an external of ca. one third of the length of the ala, a median of half of the length of the ala and an internal longer, with ca. two thirds of the length of the ala. Oesophagus with narrow, sub-cylindrical procorpus, its diameter diminishing towards the cylindrical isthmus, equaling its diameter. Basal bulb rounded, valve-plate well developed. Short cardia present, projecting into the intestine, which is simple, sub-rectilinear, narrow, its anterior region barely dilated. Walls of intestine narrow, formed by a single layer of flattened cells. Near junction with the short rectum intestinal wall becomes wider, with thicker cells. Nerve ring at level of the junction of the procorpus with the isthmus. Excretory pore ventral, ca. one third of the body width posterior to the basal bulb. Vulva a ventro-median transverse slit, its lips slightly prominent (the anterior lip more prominent than the posterior), located ca. the end of the first third of body. Body not contracted posterior to the vulva. Vagina with a muscular portion anteriorly directed, becoming less muscularized and reflexing posteriorly for a short distance before joining the uteri. Genital tract didelphic-amphidelphic. Ovaries very slender. Anterior ovary reflexed and surrounding the intestine posteriorly to its junction with the basal bulb. Posterior ovary reflexed a short distance before the junction of the intestine with the rectum. Oöcytes in single rows. Spermatheca with irregular lobes present in the posterior uterus, at the point where it reflexes anteriorly. Spermatheca absent in the anterior branch of the genital tract. Eggs ellipsoidal, shell thin and smooth. Tail very long, ca. the half of the body length, filiform, subulate, ending in a fine tip. Phasmids pore-like, lateral, located in the first portion of the tail, ca. 60 µm posterior to the anus.
Male. Body very small and comparatively more slender than that of females, its posterior end ventrally curved. Anterior end bluntly truncated. Cephalic capsule smooth, long, conoid and slightly dilated, well set-off from the rest of the body cuticle. Body cuticle finely annulated posterior to cephalic capsule. Lateral alae present, extending from the oesophageal region (at a distance of ca. 75 µm from the cephalic end) to ca. 25 µm before the level of the cloaca. Papillar structures present around the oral opening not evident. Amphids slit-like, lateral in position. Buccal cavity very short, with cuticular, irregularly folded structures projecting to the center of the lumen. Oesophagus consisting of a sub-cylindrical procorpus, with a distal dilation just posterior to the buccal cavity, diameter uniform posterior to dilation until the posterior third, where it increases slightly and diminishes again towards the junction with the isthmus. Basal bulb rounded, with valve-plate well-developed. Short cardia projecting into the intestine, simple, its anterior region not dilated. Nerve ring encircling the procorpus just anterior to junction with the isthmus. Excretory pore ventral, located at the level of the junction of the isthmus with the basal bulb. Monorchic. Distal end of the testis located at level of the junction of the isthmus with the basal bulb. Distal portion of the testis is filled with granular cells that gradually form rounded, smaller spermatocytes. The median, dilated part of the testis is filled with rod-like spermatids, with one of the ends wider. The walls of this part consist of a single layer of swollen vacuolated cells. Testis diameter decreases towards the vas deferens, lined by a single layer of cells. Ejaculatory duct narrow. Spicule and gubernaculum absent. Cloaca with the anterior lip projecting posteriorly, with its ventral side presenting randomly scattered knob-like ornamentations. One pair of long, digitiform ventral papillae located just before the projected anterior lip of cloaca. Proximal ends of these papillae are almost in contact, with their distal ends opposed. One pair of large, prominent, lateral adcloacal papillae present, its surface with curled ornamentations. Spatula-like post-cloacal projection is present, with two sensillae in the distal end. Another smaller and slender projection, also bearing two distal sensillae originating dorsal to the spatula-like structure, arising from its proximal third. Distal ends of both projections located at the same level. Tail appendage located dorsally to the spatula-like post-cloacal projection, comparatively long, filiform, ending in a fine tip.
Type locality. Chubu University Campus , Kasugai, Aichi prefecture, Japan
Type host. Parafontaria laminata (Attems, 1909) (Diplopoda: Polydesmida : Xystodesmidae ).
Site. Hindgut.
Etymology. Specific epithet dedicated to Cláudia S. L. Vicente, former postdoctoral researcher at the Department of Environmental Biology of the College of Bioscience & Biotechnology, Chubu University and specialist on bacteria-nematode-arthropod interactions.
Differential diagnosis. The species of the genus Travassosinema can be distinguished on the basis of two conspicuous female features, namely the presence/absence of lateral alae and the presence/absence of a contraction of the body width posterior to the level of the vulva, due to a notable swelling of the anterior lip of vulva. Based on such features, the closest species to T. claudiae n. sp. are T. travassosi and T. thyropygi , which as T. claudiae n. sp. also lack lateral alae and the contraction posterior to vulva. T. claudiae n. sp. differs from T. thyropygi by its shorter body (1.87–2.87 mm vs. 2.41–3.21 mm) and tail (c = 2.27–2.87 vs. 1.90–2.30). However, the eggs are slightly larger in T. claudiae n. sp. (56–69×34–44 vs. 53×39). The nerve ring in T. claudiae n. sp. is located in the procorpus-isthmus junction, instead of anterior to the basal bulb in T. thyropygi ( Hunt 1996) . T. claudiae n. sp. presents smaller eggs than T. travassosi (56–69×34–44 vs. 70×60). Furthermore, the cephalic umbraculum of T. claudiae n. sp. extends until the posterior third of the procorpus vs. the midpoint of the basal bulb in T. travassosi ( Rao 1958) .
The other species without lateral alae are T. dechambrieri , T. jaidenae and T. mirabile . From these, T. claudiae n. sp. differs by lacking the contraction of the body posterior to the level of the vulva, present in the aforementioned species. T. claudiae n. sp. has a longer body (1.87–2.87 mm vs. 1.66 ± 0.25 mm) and the vulva is more anterior (V% = 32.53–38.18 vs. 45.00 ± 3.00) than in T. dechambrieri . A similar case is T. mirabile , also with a shorter body (1.30–1.59 mm vs. 1.87–2.87 mm) and more posterior vulva (V% = 39.00–44.00 vs. 32.53–38.18) than the present species. T. claudiae n. sp. differs from T. jaidenae by having alae at the apical part of the umbraculum that correspond to the lips with widened, overlapped lobes that obscure the interlabia. In T. jaidenae such lobes are not developed and the apical part of the umbraculum alae of the lips do not contact each other ( Jex et al. 2005). As in T. dechambrieri and T. mirabile , the vulva of T. claudiae n. sp. is more anteriorly located than in T. jaidenae (V% = 32.53–38.18 vs. 37.00–45.00).
In addition to T. claudiae n. sp., males have been described only in T. travassosi , T. mirabile and T. dalei . The latter males resemble those of T. claudiae n. sp. by having a similar body length (515–733 µm vs. 584–693 µm), form of the oesophagus, absence of spicule and gubernaculum and presence of a tail filament. The structures around the cloaca are also similar, with the characteristic mid-ventral, digitiform pre-cloacal pair of papillae, a large pair of papillae and the anterior lip of the cloaca ornamented and, having a spatula-like post-cloacal projection. Both species differ in having a slightly shorter oesophagus length in the males of T. claudiae n. sp. (b = 3.38–4.01 vs. 2.60–3.60). The position and shape of the large ornamented papillae is different in these species: more adcloacal and being more prominent, with curled ornamentations in opposition to the post-cloacal, and less prominent, with tuberculate ornamentations of T. dalei . The minute pair of post-cloacal papillae in the tail filament of T. dalei was not observed in T. claudiae n. sp. Also, males of T. dalei lack lateral alae. Alae are present in T. claudiae n. sp., extending from the oesophageal region to a short distance anterior to the level of the cloaca.
Males of T. claudiae n. sp. can be easily differentiated from T. mirabile and T. travassosi by lacking a spicule and having a long, filiform tail filament. Both latter species present a single rod-like spicule and the tail filament is very short and rounded in T. travassosi and thick in T. mirabile . Moreover, the number and arrangement of copulatory papillae is different. Males of T. mirabile present numerous papillae including eight pre-cloacal pairs (six sub-ventral and two sub-dorsal), one pair adanal before the cloacal opening, another pair behind the cloacal opening and two post-cloacal pairs on a terminal tail protrusion. On the other hand, males of T. travassosi have two pre-cloacal and one adanal pairs. Spiridonov and Cribb (2012) discussed the possibility of a misidentification of the males of T. mirabile and T. travassosi , considering the differences among both species with those of T. dalei .
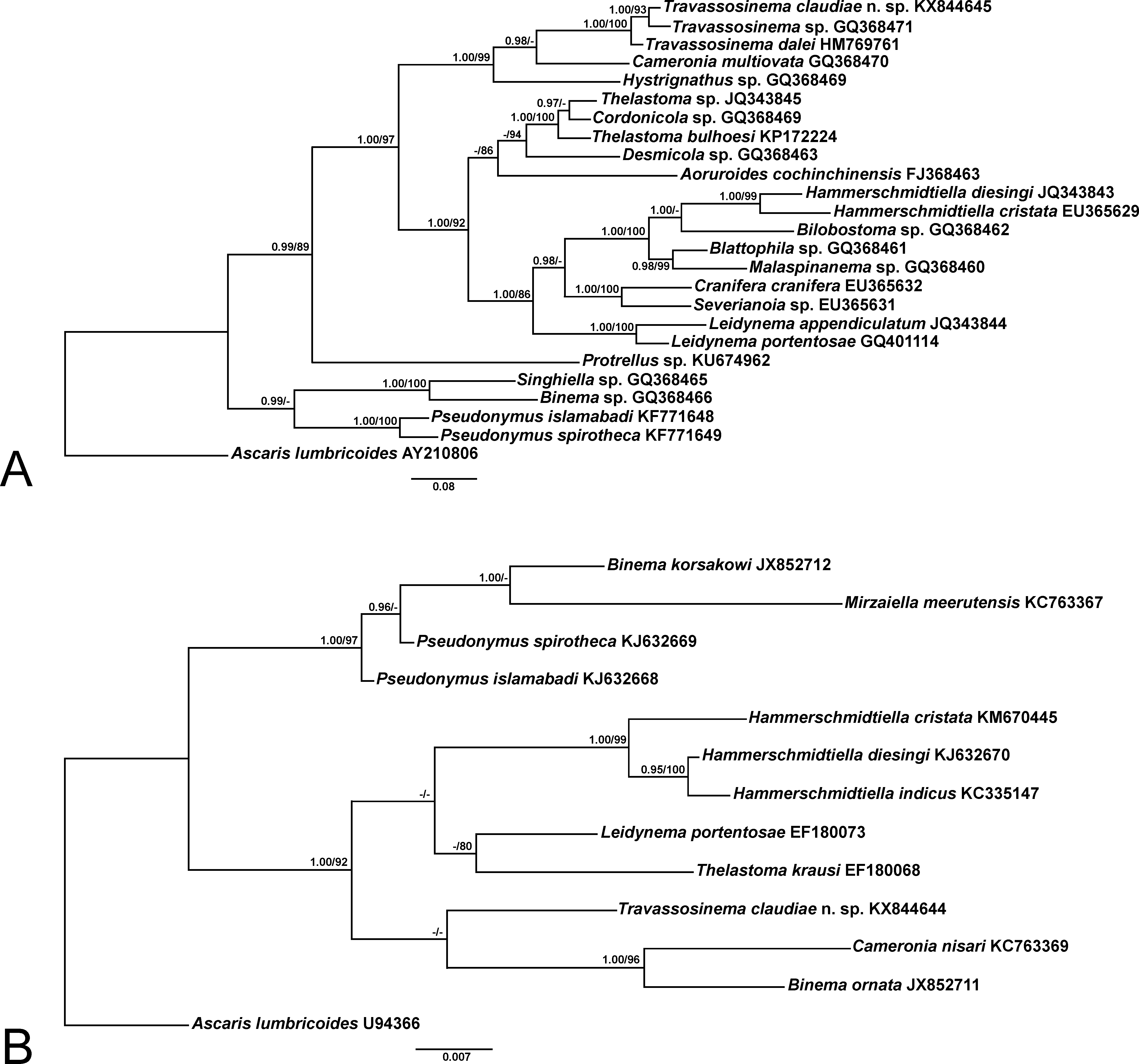
Phylogeny. The topology for both, ML and BI trees was identical and only the BI trees are shown ( Fig. 5 View FIGURE 5 ). In addition to T. claudiae n. sp. only two D2-D3 LSU partial sequences of Travassosinema are available: Travassosinema sp. GQ368471 View Materials , from a Vietnamese millipede identified as Thyropygus sp. ( Spiridonov & Guzeeva 2009) and T. dalei HM769761 View Materials from an Australian scarabeid beetle larva ( Spiridonov & Cribb 2012). T. claudiae n. sp. differs from both species by 28 bp in an alignment of 725 bp. The three species form a well-supported clade, the new species being closer to Travassosinema sp. also from a millipede and with T. dalei basal in position. Coinciding with Spiridonov & Guzeeva (2009) Cameronia Basir, 1948 constitutes the sister group of Travassosinema and Hystrignathus sp. is basal in position in the well-supported clade formed by Travassosinema + Cameronia + Hystrignathus . For the 18S SSU (where the new species constitutes the first sequence of Travassosinema ) T. claudiae n. sp. is located as the sister taxon to Cameronia nisari (Parveen & Jairajpuri, 1985) and Binema ornata Travassos, 1925 .
For both markers it is striking that genera currently considered as belonging to the family Travassosinematidae are located in different and phylogenetically distant clades. In the case of the D2-D 3 28S LSU the other travassosinematids included in the analysis, Binema sp. and Singhiella sp., form a monophyletic clade with two species of Pseudonymus Diesing, 1857 (Pseudonymidae) . Similar to the 28S LSU, in the 18S SSU analysis two travassosinematids: Binema korsakowi (Sergiev, 1923) and Mirzaiella meerutensis Singh & Malti, 2003 form a clade with both Pseudonymus species. Genera Binema Travassos, 1925 ; Mirzaiella Basir, 1942 and Singhiella Rao, 1958 consist of nematodes from mole crickets whereas Pseudonymus is present in aquatic beetles of the family Hydrophilidae . The proximity of these taxa, which belong to different families, could be justified by sharing some characteristic morphological features, namely the presence of polar filaments in the eggs and males with more than four pairs of copulatory papillae.
The present molecular data contradict the results of Adamson & Van Waerebeke (1992), who placed the umbraculum-bearing genera Indiana and Pulchrocephala and the parasites from mole crickets along with Travassosinema within the family Travassosinematidae . According to his phylogenetic analysis of these taxa, Adamson (1987) found that the three umbraculum-bearing genera formed a monophyletic clade. However, the author disagreed with the criteria of Rao (1958) about placing such three genera within Travassosinematidae , since it would create contradictions for the placement of the other genera from mole crickets.
The genera Indiana and Pulchrocephala are more similar to the other genera of parasites from mole crickets, namely Binema , Chitwoodiella Basir, 1948 ; Isobinema Rao, 1958 ; Mirzaiella , Mohibiella Farooqui, 1970 ; Pteronemella Rao, 1958 and Singhiella . All these, excepting Pteronemella where the females are unknown, have in common the presence of eggs with polar filaments, a synapomorphy shared only with some genera of Pseudonymidae . The males lack of a caudal filament, with the exception of Binema and Isobinema , and the number of copulatory papillae is more than four. The genus Travassosinema differs from the latter genera by lacking polar filaments in the eggs. Considering the possibility that the males described as conspecific with T. mirabile and T. travassosi belong to a different genus ( Spiridonov & Cribb 2012) the males of T. claudiae n. sp. and T. dalei present a caudal filament and less than four pairs of copulatory papillae.
A main reason for the arrangement of Travassosinema along with Indiana and Pulchrocephala within Travassosinematidae was to consider the cephalic umbraculum as a homologous structure for the three genera ( Adamson & Van Waerebeke 1992). Adamson (1987) explained the possible evolution for the cephalic umbraculum, being Travassosinema a more primitive pattern than the other umbraculum-bearing genera. The author also stated that there are elements of the umbraculum that are homologous for the three genera. It is evident that morphologically, the cephalic umbraculum of Travassosinema is more different than those of Indiana and Pulchrocephala , with six elements arranged as an anterior circle of one dorsal and two sub-lateral alae derived from the labia and a posterior circle of one ventral and two sub-lateral alae derived from the interlabia. The other genera present 12 elements in the umbraculum. Indiana presents an anterior circle of six elements, one dorsal, one ventral and four sub-lateral plus a posterior circle of six elements located between the alae of the anterior circle, two lateral, two sub-dorsal and two sub-ventral ( Adamson & Van Waerebeke 1987). Hunt (1983) had different criteria and considered the elements of the first circle as lateral and sub-median. Pulchrocephala also presents 12 elements, the anterior circle with six hood-like expansions, one dorsal, one ventral and four sub-lateral and a posterior circle with six alae, in the same position as the anterior one ( Hunt 1981).
Considering the current results, the monophyletic composition of the family Travassosinematidae with the genus Travassosinema along with the other genera of thelastomatoids from mole crickets is questionable. The morphological differences between the Travassosinema species with the other travassosinematid genera, as well as their specificity for groups of hosts other than mole crickets point towards a different taxonomic placement. The inclusion of molecular data and more detailed morphological studies, especially on the remaining umbraculumbearing genera Indiana and Pulchrocephala as well as other travassosinematid genera is needed in order to clarify the latter statements.

| CZACC |
Coleccion Zoologia, Academia de Ciencias de Cuba |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Oxyuridomorpha |
|
Family |
|
|
Genus |