
Urolibellula eocenica, Zeiri, Asma, Nel, Andre & Garrouste, Romain, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4032.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:EC2FE508-40E1-4464-868D-E2ED0BC56960 |
|
DOI |
https://doi.org/10.5281/zenodo.6111617 |
|
persistent identifier |
https://treatment.plazi.org/id/0B5287CD-4801-FFF9-FF42-9984FAA7A085 |
|
treatment provided by |
Plazi |
|
scientific name |
Urolibellula eocenica |
| status |
sp. nov. |
Urolibellula eocenica sp. n.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Material. Holotype specimen without number, labelled “Dragonfly, Anisoptera gen. Corduliidae ”, USNM (United State National Museum).
Age and outcrop. Eocene, Green River Formation, Parachute Creek Member, 43–47 Myrs, Wighton, Garfiel County, Colorado.
Diagnosis. As for the genus.
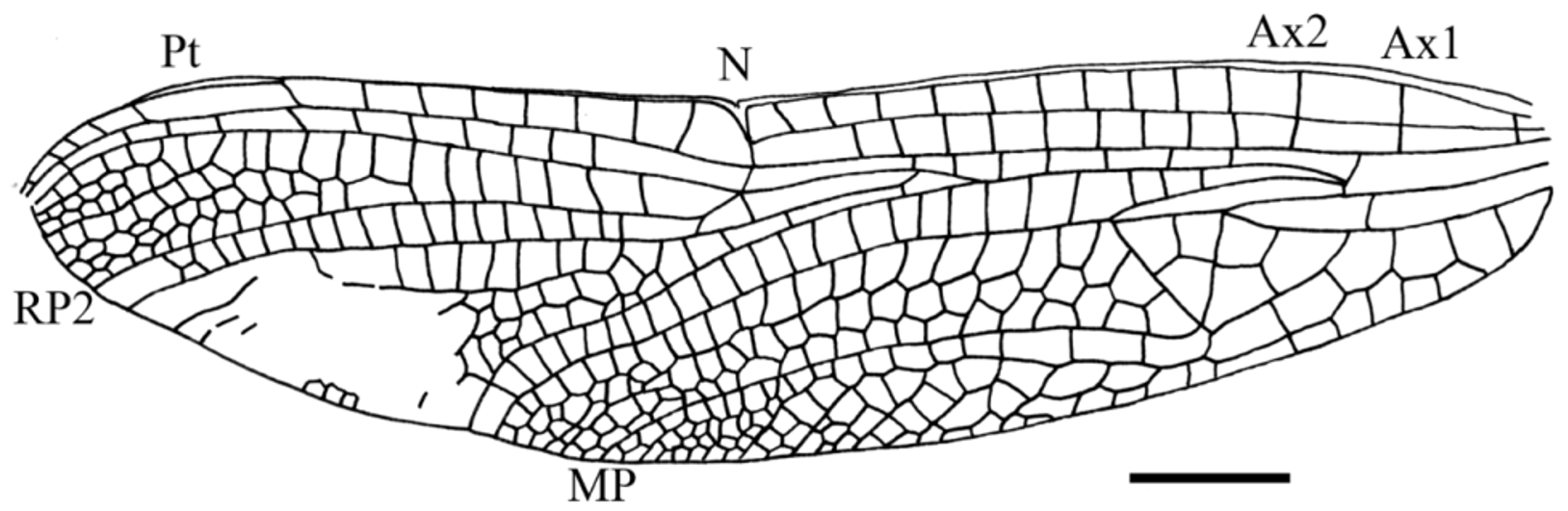
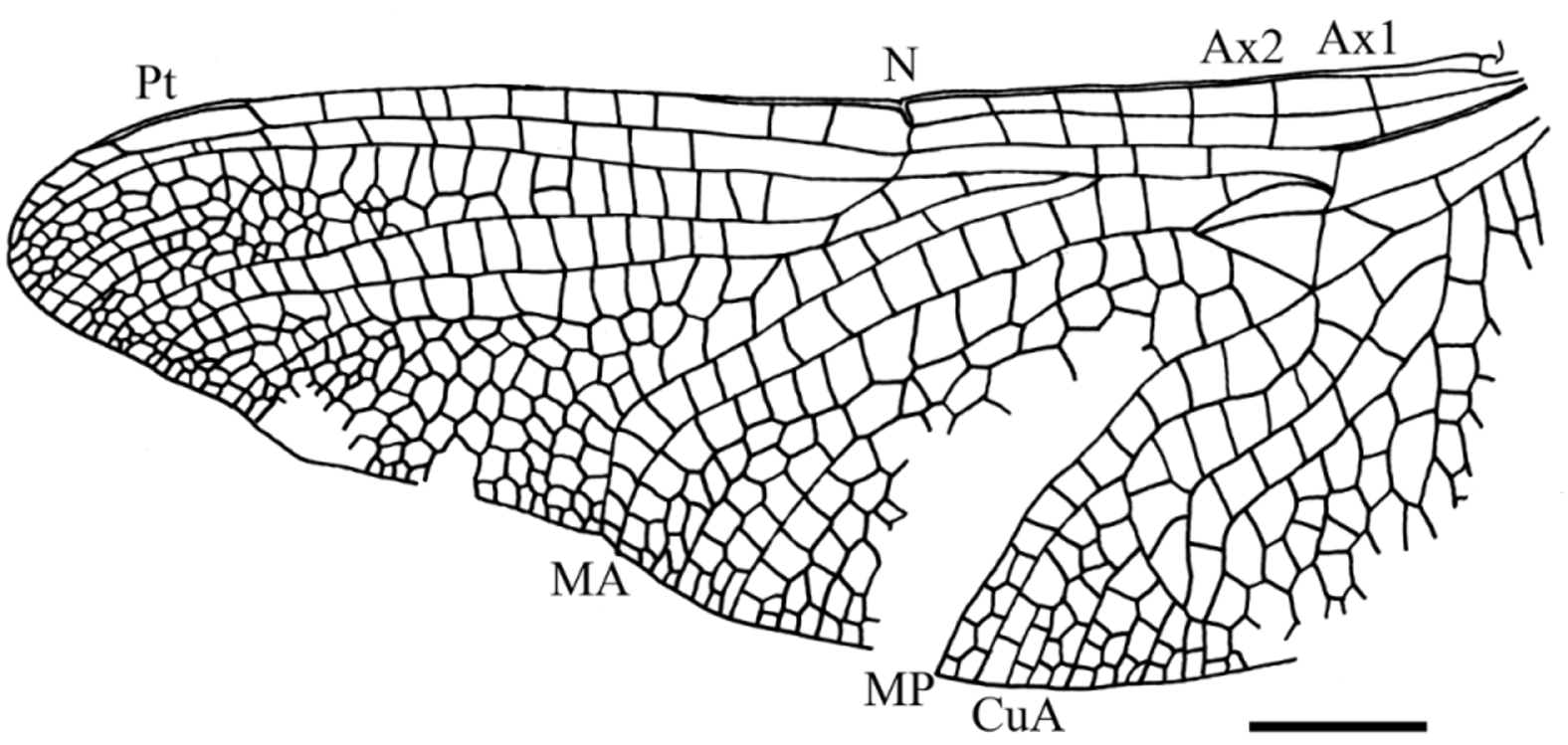
Description. Two forewings, one hind wing, and three legs attached to fragments of the thorax. Wings hyaline, pterostigmata dark brown.
Forewing length 38.0 mm; width at nodus 8.0 mm; distance from base to nodus 20.0 mm; nodus situated distinctly closer to apex than to base; distance from nodus to pterostigma 11.0 mm; distance from base to arculus 6.0 mm; primary antenodal crossveins Axl and Ax2 aligned but not stronger than other 10 secondary antenodals (all complete except most distal one, but not well aligned with antenodals of second row between ScP and RA); Axl 2.0 mm basal of arculus and Ax2 5.0 mm distal of Axl, a long “cordulegastrid gap” and a gap directly distal of arculus; seven postnodal crossveins between nodus and pterostigma not aligned with six postsubnodal crossveins; a “libellulid gap” directly distal of subnodus concerning one postnodal; pterostigma 4.0 mm long and 1.0 mm wide; pterostigma with a basal brace not aligned with its basal side, distally displaced and weakly oblique; pterostigma covering two cells; arculus nearly totally straight and aligned with basal side of discoidal triangle; origins of RP and MA (sectors of arculus) separated at their extreme bases in arculus; free portion of RP +MA longer than posterior crossvein in arculus; hypertriangle 3.8 mm long and 1.0 mm wide, free; its costal side (MA) weakly curved; discoidal triangle not transverse but broad, divided into three cells by crossveins touching in one point; length of basal side of discoidal triangle 3 mm, length of costal side 2.8 mm, length of distal side MAb 4 mm; MAb straight; pseudo-anal vein PsA distinct; subdiscoidal triangle three-celled; basal space free; submedian space only crossed by CuP; anal area max. 2.0 mm wide with two rows of cells; cubito-anal area max. 4.5 mm wide with up to five rows of cells; MP weakly curved and ending just distal of nodus; area between CuA and MP not widened; basal part of postdiscoidal area with 2–3 rows of cells; postdiscoidal area not widened distally (width near discoidal triangle 2.5 mm; width at hind margin 2.3 mm); Mspl nearly straight, ending at MAa, with one row of cells between it and MAa; MAa weakly curved; RP 3/4 and MA weakly undulate and closely parallel with only one row of cells between them up to hind margin; first branching of RP 6.1 mm basal of subnodus; IR2 originating on RPl/ 2; RP 2 aligned with subnodus; only one lestine oblique vein “ O ” between RP 2 and IR2, 0.5 mm and one cell distal of subnodus; presence of a post-oblique-vein gap; second crossvein between RP 1 and RP 2 developed as a “libellulid oblique vein;” only one bridge crossvein between RP 2 and IR2 basal of subnodus; area between RP 2 and IR2 distally widened, but only one row of cells between these veins, except near hind margin (two rows of cells); a well-defined Rspl with one row of cells between it and IR2; RP 2 weakly undulate; RPl and RP 2 basally parallel with only one row of cells between them, but about 1.0 mm basal of pterostigma, area between these veins widening progressively; a well-defined pseudo-IRl originating on RPl beneath pterostigma; three rows of cells between pseudo-IRl and RPl and between pseudo-IRl and RP 2; no apical supplement (strong concave supplementary sector developed between RP 1 and RP 2).
Hindwing: length 37.0 mm; width at nodus 15.0 mm; distance from base to nodus 15.0 mm; nodus situated distinctly closer to base than to apex; distance from nodus to pterostigma 15.5 mm; distance from base to arculus 5.5 mm; primary antenodal crossveins Axl and Ax2 aligned but not stronger than other five antenodals (all complete); Axl 3 mm basal of arculus and Ax2 is 5.5 mm distal of Axl; only four antesubnodal crossveins, with a long “cordulegastrid gap” and a gap directly distal of arculus; 10 postnodal crossveins between nodus and pterostigma not aligned with eight postsubnodal crossveins; a “libellulid gap” directly distal of subnodus concerning two postnodals; pterostigma 3.5 mm long and 1.0 mm wide; pterostigma with a basal brace not aligned with its basal side, distally displaced and weakly oblique; pterostigma covering two cells; arculus nearly totally straight and aligned with basal side of discoidal triangle; origins of RP and MA (sectors of arculus) separated at their extreme bases in arculus; free portion of RP +MA longer than posterior crossvein in arculus; hypertriangle 3.0 mm long and 0.75 mm wide, free; its costal side (MA) distinctly curved; discoidal triangle not transverse but broad, and free; length of basal side of discoidal triangle 1.8 mm, length of costal side 3.0 mm, length of distal side MAb 3.0 mm; MAb straight; pseudo-anal vein PsA distinct; subdiscoidal triangle unicellular; basal space free; submedian space only crossed by CuP; anal area max. 12.5 mm wide with about ten rows of cells; cubito-anal area max. 8.0 mm wide with up to six rows of cells; CuA strongly sigmoidally curved and with a strong CuAb; anal loop long and narrow, posteriorly closed and with a concave well-defined midrib with a distinct toe; MP strongly curved and ending just basal of nodus; area between CuA and MP not widened; basal part of postdiscoidal area with only two rows of cells; postdiscoidal area strongly widened distally (width near discoidal triangle 3.5 mm; width at hind margin 7.0 mm); a Mspl nearly straight ending on MAa, with one row of cells between it and MAa; MAa weakly curved but with a strong bend near posterior wing margin; RP 3/4 and MA not undulate and closely parallel with only one row of cells between them up to hind margin; first branching of RP 5.0 mm basal of subnodus; IR2 originating on RPl/2; RP 2 aligned with subnodus; only one lestine oblique vein “ O ” between RP 2 and IR2, 0.5 mm and one cell distal of subnodus; presence of a post-oblique-vein gap; second crossvein between RP 1 and RP 2 developed as a “libellulid oblique vein;” only one bridge crossvein between RP 2 and IR2 basal of subnodus; area between RP 2 and IR2 distally widened, but only one row of cells between these veins, except near hind margin (two rows of cells); a well-defined Rspl with one row of cells between it and IR2; RP 2 weakly undulate; RPl and RP 2 basally parallel with only one row of cells between them, but about 1.5 mm basal of pterostigma, area between these veins widening progressively; a well-defined pseudo-IRl originating on RPl beneath pterostigma; three rows of cells between pseudo-IRl and RPl and between pseudo-IRl and RP 2; no apical supplement (strong concave supplementary sector developed between RP 1 and RP 2); basal hind margin not well preserved so that the presence of an anal angle or an anal triangle cannot be ascertained; no membranule visible.
Discussion. The presence of a post-oblique-vein gap is a specialized character present in the Macromiidae and most taxa of Haplohamulida (sensu Bechly 2005). The presence of a well-defined Mspl, a well-developed, elongate, boot-shaped anal loop with a distinct toe and a distinct Cuspl) is a synapomorphy of the clade Italoansida Bechly, 1996 ( Corduliidae Selys in Selys & Hagen, 1850, + Anauriculida Bechly, 1996). The following putative synapomorphies support a position in the Anauriculida (= Hemicorduliidae Bechly, 1996 , + Libellulida Bechly, 1996) for Urolibellula gen. n.: two primary antenodal brackets Ax1 and Ax2 less distinct; besides anal crossing (= CuP) no cubito-anal crossveins present in hindwing (convergent to Pseudocordulia , Austrocordulia , Neophya , Hesperocordulia ; maybe synapomorphic with some Corduliidae ; reversed in some derived libellulids) because of reduction of the basal side of the hindwing subdiscoidal triangle; characteristic curved shape of Rspl that is distally “rejoining” IR2; Mspl very distinct and curved, distally “rejoining” the MA. Urolibellula differs from the Hemicorduliidae ( Hemicordulia Selys, 1870 ) in the most distal antenodal crossvein incomplete and oblique in forewing, presence of a one-celled subdiscoidal triangle in hindwing and discoidal triangle of forewing with three cells not separated by parallel crossveins. Urolibellula shares with the Libellulida (= Libellulidae Leach, 1815 , + Urothemistidae Lieftinck, 1954 ) the following synapomorphies (note that in the phylogeny of the higher Libelluloidea proposed by Ware et al. (2007), the Urothemistidae are considered as a subfamily Urothemistinae of the Libellulidae but still falls as sister group of the remaining Libellulidae , in the same position as in Bechly (1996)): the two primary antenodal brackets Ax1 and Ax2 are very indistinct in forewings and indistinguishable from secondaries in hindwings (even if some secondary antenodals of forewing are not well aligned); anal loop reaching hind margin of hindwing or nearly so (well-developed “toe”) ( Figs. 4–5 View FIGURE 4 View FIGURE 5 ). Bechly (1996) proposed a further character for this clade, i.e. the second crossvein between RP 1 and RP 2 is developed as a “libellulid oblique vein.” This character is not very pronounced in Urolibellula , as this vein is only weakly oblique, but some modern Libellulidae (e.g. Libellula foliata Kirby, 1890 ) also have this vein not very oblique. Urolibellula differs from the Urothemistidae in the libellulid oblique vein not very oblique, most distal antenodal incomplete in forewing, presence of a subdiscoidal triangle in hindwing, and forewing discoidal triangle much broader (see Ris 1909 – 1913). Urolibellula shares with the Libellulidae the following apomorphies: most distal antenodal between costal margin and ScP developed as antenodal oblique vein, with suppression of corresponding antenodal between ScP and RA in forewing; all antenodal crossveins developed as antenodal brackets, therefore the two primaries Ax1 and Ax2 completely indistinguishable from the secondaries in hindwings. This character is also present in the forewings of many Libellulidae but not in all (e.g. Hydrobasileus Kirby, 1889 , or the oldest libellulid Palaeolibellula). Nevertheless, Urolibellula lacks two other putative apomorphies of the Libellulidae , i.e. its sectors of arculus are not stalked and it has no apical supplement (strong concave supplementary sector developed between RP 1 and RP 2). Its forewing discoidal triangle is clearly broader than in any modern Libellulidae and divided into three smaller cells by crossveins not longitudinally disposed. Its subdiscoidal triangle and triangle are both very large with approximatively the same sizes. The modern Libellulidae with triangle and subdiscoidal triangle of similar sizes have a narrow triangle and a subdiscoidal triangle quite smaller than in Urolibellula . The forewing discoidal triangle and subdiscoidal triangle of Urolibellula reminds one of some “corduliids” (e.g. Epitheca Charpentier, 1825 ) (see Martin 1914), even of some Petalurida ( Protolindenia Deichmüller, 1886 ) or Aeshnoptera (Paramesuropetala Bechly et al., 2001) ( Nel et al. 1998; Bechly et al. 2001). Thus this character is clearly subject to convergencies. In conclusion, the Urolibellulidae would fall in a “basal” position as probable sister group of the Libellulidae (sensu Bechly 1996, 2014), the Urothemistidae being the sister group of the (Urolibellulidae + Libellulidae ). The presence of a representative of the crown group of the Libellulidae in the Late Cretaceous shows that the Urolibellulidae were certainly also present in the Late Mesozoic. The stem lineage of the Libellulidae was probably diversifying during the Late Cretaceous-Eocene even if the fossil record of this clade is very scarce for this period.
Remarks. Cockerell (1921) described a fossil abdomen from the Eocene Green River Formation under the name Stenogomphus scudderi . The ‘libelluloid’ genus Stenogomphus Scudder, 1892 is based on a forewing from the Earliest Oligocene Florissant Formation ( Scudder 1892; Ris 1910; Nel & Paicheler 1994). There is no argument supporting the inclusion of S. scudderi in the genus Stenogomphus . S. carletoni Scudder, 1892 differs from our fossil in the complete last antenodal crossvein, and the second crossvein between RP 1 and RP 2 not oblique.



| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.