
Pristimantis ardilae, Acevedo & Armesto & Palma, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4750.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:5FE0773B-3C35-4D30-8CE7-F1691ABA5F83 |
|
DOI |
https://doi.org/10.5281/zenodo.3717962 |
|
persistent identifier |
https://treatment.plazi.org/id/0D4A4E06-FFEF-A73A-FF08-FB85FA1274BA |
|
treatment provided by |
Plazi |
|
scientific name |
Pristimantis ardilae |
| status |
sp. nov. |
Pristimantis ardilae , sp. nov.
( Figure 4A View FIGURE 4 )
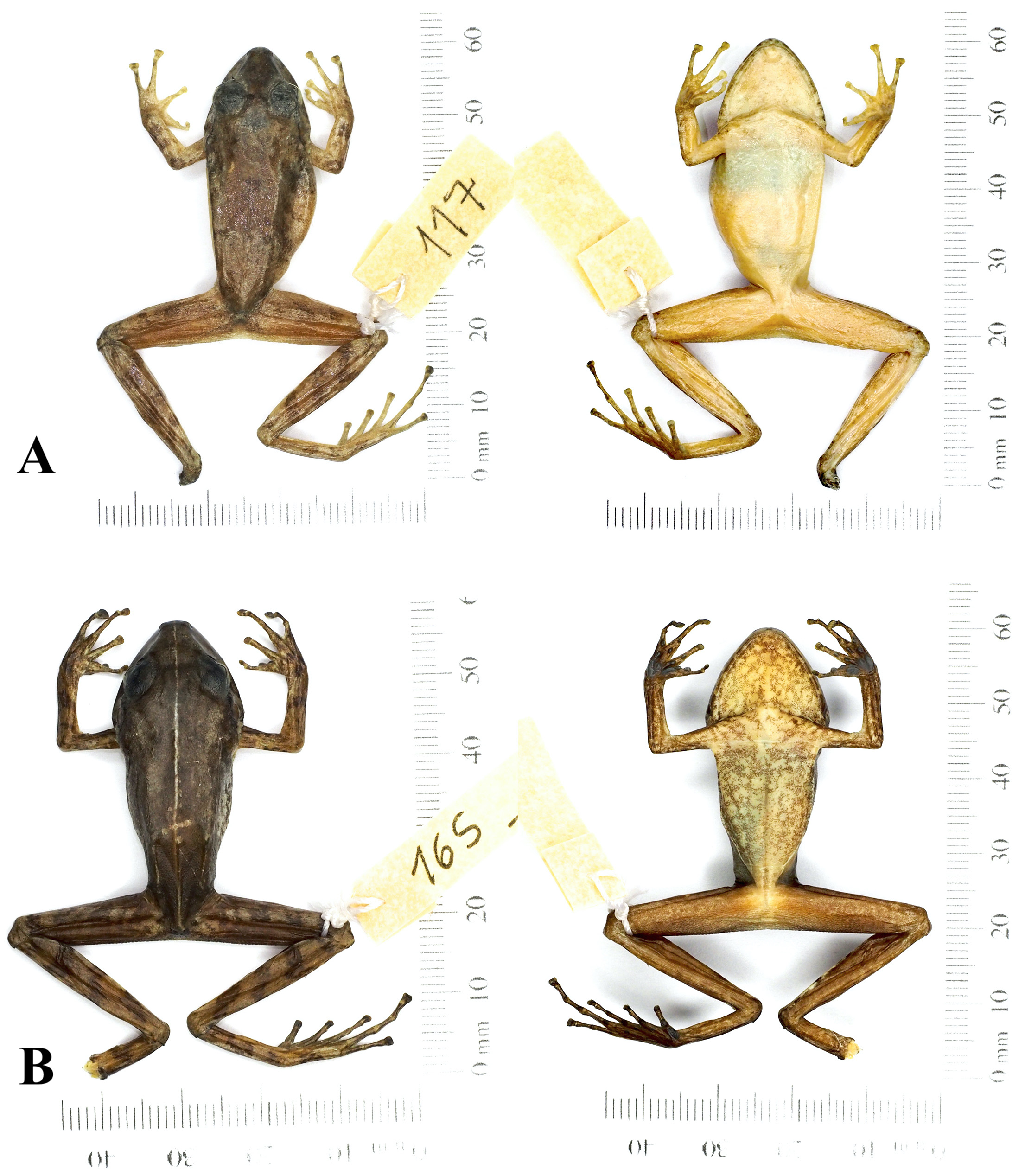
Holotype. An adult female ( MCNUP-H 117 , Figure 5A View FIGURE 5 ), collected on September 10, 2010, by Aldemar A. Acevedo, Karen Silva, and Rosmery Franco in the locality of San Antonio (7.162172 –72.25254; 612 m. a.s.l.), municipality of Toledo in the buffer zone of the Tamá National Natural Park ( PNNT). GoogleMaps
Paratypes. Two adult females ( MCNUP-H 121 and 157), and two adult males ( MCNUP-H 166 and 253) collected at the type locality on the same date of the holotype. One adult female ( MCNUP-H 163 ) , and two adult females ( MCNUP-H 673 and 247), Collected on May 3, 2013, by Aldemar A. Acevedo in the low area of the PNNT, La Vega (7.13612 –72.1843; 465 m. a.s.l.), municipality of Toledo in the buffer zone of the Tamá National Natural Park ( PNNT) GoogleMaps .
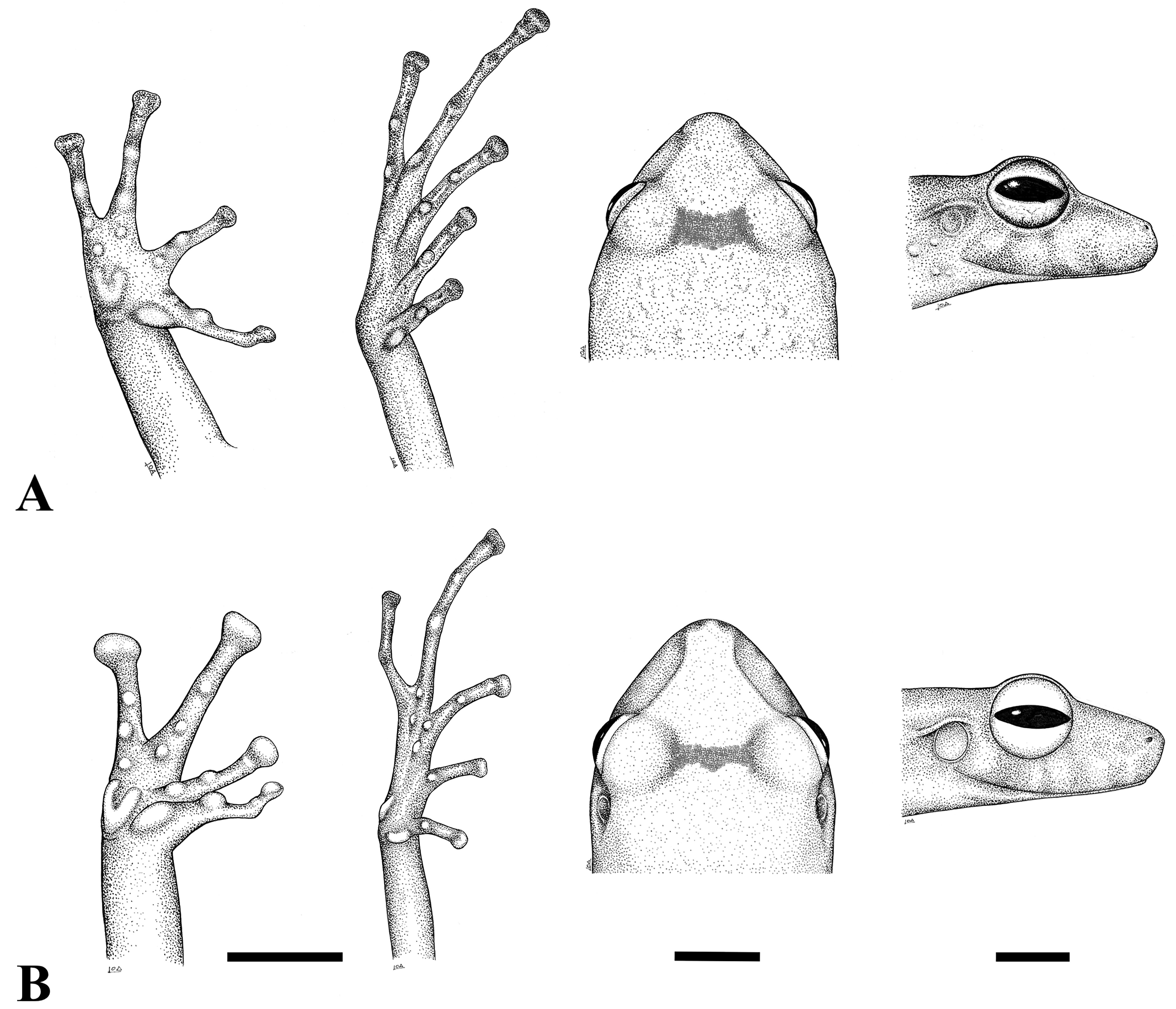
Definition and diagnosis. Pristimantis ardilae , sp. nov. is assigned to the genus Pristimantis based on molecular phylogenetic relationships ( Figure 2 View FIGURE 2 ) and its morphological characteristics ( Lynch & Duellman 1997; Duellman & Lehr 2009). We assign the new species to the P. conspicillatus species group based on position obtained in our phylogenetic analyses ( Figure 2 View FIGURE 2 ), and following Lynch & Duellman (1997) and Padial & De la Riva (2009) for having: (1) skin on dorsum shagreen; venter smooth to finely areolate whitish or yellowish coloration, and some black spots on the throat; short dorsolateral folds in scapular region. (2) tympanum distinct, round, ~1/2–1/3 of the eye diameter; tympanic membrane and annulus present; supratympanic fold evident. (3) snout subacuminate in a dorsal view; rounded in profile ( Figure 6A View FIGURE 6 ). (4) canthus rostralis defined, straight in dorsal view, slightly rounded in cross section; loreal region weakly concave. Nostrils slightly distinct, directed laterally. (5) upper eyelid without tubercles; cranial crests absent. (6) choanae small, round; dentigerous processes of vomers evident, with 5–6 teeth on the posterior margin; 12–16 premaxillary teeth. (7) vocal slits and nuptial pads present. (8) ED: 89% (females) and 96% (males) of the END. IOD: 81 % (females) and 78 % (males) of the ED. (9) THL ~51% (females) and 46% (males) of the SVL. TL ~ 58% (females) and 56% (males) of the SVL. FL ~ 47 % (females) and 46 % (males) of the SVL. HW ~ 37% of the SVL in both females and males. (10) ulnar tubercles absent. (11) relative lenght of fingers: II ≤ I < IV < III ( Figure 6A View FIGURE 6 ). Subarticular tubercles are round and upraised. The palmar tubercle is bifid, shaped similar to a heart; supernumerary tubercles is rounded and pronounced below the proximal subarticular tubercle of FIV; the thenar tubercle 80% of palmar tubercle. Finger fringes absent. FIII ~30% longer than FI. Disks on FIII–IV moderately expanded (state 2 according to Grant et al. 2006); disks on FI–II weakly expanded (state 1 according to Grant et al. 2006). (12) inner metatarsal tubercle ovoid; outer metatarsal tubercle small; tarsus without fold; relative lenght of toes: I < II < III < V < IV; Toe disks elliptical; Toe webbing absent ( Figure 6A View FIGURE 6 ). (13) Pristimantis ardilae sp. nov. is apparently sexually dimorphic in size, with a mean SVL of 27.5 mm (n = 4) for males and 38.3 mm (n = 4) for females ( Table 2).
Comparisons with other species. Pristimantis ardilae sp. nov. can be further distinguished from other Pristimantis species from the northeastern region of Colombia, the Venezuelan Andes, and phylogenetically related species of the P. conspicillatus species group by the combination of characters summarized in Table 3.
Of the 33 species that are part of the P. conspicillatus species group, only P. carranguerorum , and P. medemi (Lynch 1994) are found in the Eastern Cordillera of Colombia, the remaining species are found in distant biogeographic areas (Amazon and Cerrado, the Guianas and in the Atlantic Forest). Therefore, the comparison was based on Pristimantis species present in surrounding areas of the Northeast of Colombia and the Andes of Venezuela. The character state of the compared species is between parentheses. Pristimantis ardilae sp. nov. is distinguished from P. carranguerorum for having the first finger slightly longer than second (Finger I < II); fingers and toes without lateral fringes or keels (fingers and toes bearing fleshy lateral keels); Throat weakly spotted (heavily marble with brown). Tarsal fold absent (presente); P. ardilae sp. nov. is smaller (male SVL: 25.1–29.1, female SVL: 36.3–39.7) than P. carranguerorum (male SVL: 32.7 and female SVL: 41.7–50.7). Pristimantis medemi is superficially similar to P. ardilae sp. nov. but differ in the states of the following characters: short dorsolateral folds in scapular region (absent); fingers and toes without lateral keels (fingers and toes bearing lateral keels). Pristimantis ardilae sp. nov. has no marks on the groin and hidden surfaces of the hind limbs (present in P. melanoproctus ( Rivero 1984 “1982”), P. mondolfii , P. anolirex ). It is a larger species (with a maximum SVL of 39.73 mm females; 29.1 mm males) than species that are geographically and phylogenetically close, ( Table 3). Pristimantis ardilae sp. nov. does not have cranial crests (present in P. anolirex ). Its first finger is longer than the second (the first finger is shorter than or about the same size as the second in P. anolirex , P. mondolfi , P. melanoproctus ). Inner and outer edges of tarsus lacking tubercles (present in P. tubernasus ( Rivero 1984 “1982”), P. anolirex , P. nicefori ).
Description of the Holotype. SVL: 39.51 mm; upper eyelid tubercles absent in preservative (slightly visible in life); HL 38% of the SVL; HW 37% of the SVL; snout subacuminate in a dorsal view and rounded in profile; END 34% of the HW; tongue longer than wide. ED slightly shorter (88%) than the END; canthus rostralis distinguishable, straight in dorsal view and slightly rounded in cross-section; IND 70% of the END; IOD 96% of the IND. Cranial crests absent; supratympanic fold evident, tympanic annulus and tympanic membrane present, TD 34% of the ED; distance tympanum-eye: 2.96 mm. TD 9.8% of the HL. Choanae small, round; dentigerous processes of vomers with 6 teeth. Premaxilla with 16 teeth. Ulnar tubercles absent. Fingers and toes without lateral fringes. FI> FII; disks on FIII and FIV moderately expanded (state 2 according to Grant et al. 2006); relative size of the yuxtaposed fingers: II <I <IV <III. Subarticular tubercles round and upraised, the largest one on FI. Palmar tubercle bifid; supernumerary tubercles distinct, protruding; thenar tubercle 80% the size of palmar tubercle. Tarsal and calcar tubercles absent. Relative size of toes: I <II <III <V <IV; toe disks elliptical; toe webbing absent. LHU 66 % of the FOL.
Color in life. The dorsum usually intergrades yellowish brown tones against a predominantly dark brown background. Head lighter cream and copper color than the dorsal part, with a large, dark, nearly triangular, interorbital spot. The upper lip has pale stripes. The flanks are predominantly lighter, often crossed by oblique brown bands, more evident toward the inguinal area. Ventral surface light cream; throat speckled with small dark brown spots. Upper arms and forearms cream color with brown to reddish transverse bands. The inguinal area is lighter with slightly mottled with cream.
Color in preservative. Specimens fixed with 10% formalin and stored in alcohol at 75% presented a dorsum with a dark brown coloration, with the head darker than the body, the lips maintained a pattern of light and brown stripes, although less defined than in life.The ventral part maintained a clear cream coloration, with a weak pattern towards the neck identifiable by small dark brown mottles. Slightly creamy extremities, but with a predominance of a dark copper color, the arms and hind limbs displayed weak traces of dark brown stripes.
Variation. Pristimantis ardilae sp. nov. exhibits sexual dimorphism in body size with adult females reaching 39.7 mm SVL (MCNUP-H 163), adult males up to 29.1 mm SVL (MCNUP-H 253). In addition, the texture of skin dorsally is roughened and is bedecked with scattered tubercles, unlike females, whose back is usually smooth. Dorsal color pattern also differs between sexes; females tend to have a more uniform cream background on the dorsal surface and light copper stripes arranged towards the sides of the body. In contrast, the males typically have a copper background on the dorsal with the dorsolateral folds with a strong reddish marking that extended from posterior edge of the upper eyelid to the sacrum.
Distribution and natural history. Pristimantis ardilae sp. nov. is only known from two localities: the settlements of San Antonio and La Vega, 9–11 km from the road Toledo-Saravena, near the Margua river. These two localities correspond to tropical humid forest and riparian zones of the lowlands (400 and 700 m. a.s.l.) from the westen flank of the Eastern Cordillera, in the Tamá National Natural Park and its buffer zone, department Norte de Santander, Colombia ( Figure 1 View FIGURE 1 ). The individuals of P. ardilae sp. nov. were found among the riparian zone perched at a height between 1 and 2.5 meters from the ground. The highest activity was recorded between 8 and 11 at night. The species was found sympatrically with other amphibians such as P. bowara sp. nov., Rhaebo glaberrimus (Günther) , Caecilia subnigricans (Dunn) , Rulyrana flavopunctata (Lynch & Duellman) and Allobates algorei (Barrio- Amorós & Santos).
Etymology. The specific epithet refers to the professor and herpetologist María Cristina Ardila-Robayo (15 February 1947 – 24 November 2017), who was a pioneer of herpetological studies in Colombia and contributed significantly to the knowledge of the diversity of Colombian amphibians, describing 15 species of Pristimantis frogs.



| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |