
Tetragonuridae
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlab058 |
|
DOI |
https://doi.org/10.5281/zenodo.6771677 |
|
persistent identifier |
https://treatment.plazi.org/id/0E16878B-FFF4-FFC0-FCFB-F8B4FDF1FB5F |
|
treatment provided by |
Plazi |
|
scientific name |
Tetragonuridae |
| status |
|
Tetragonuridae View in CoL View at ENA
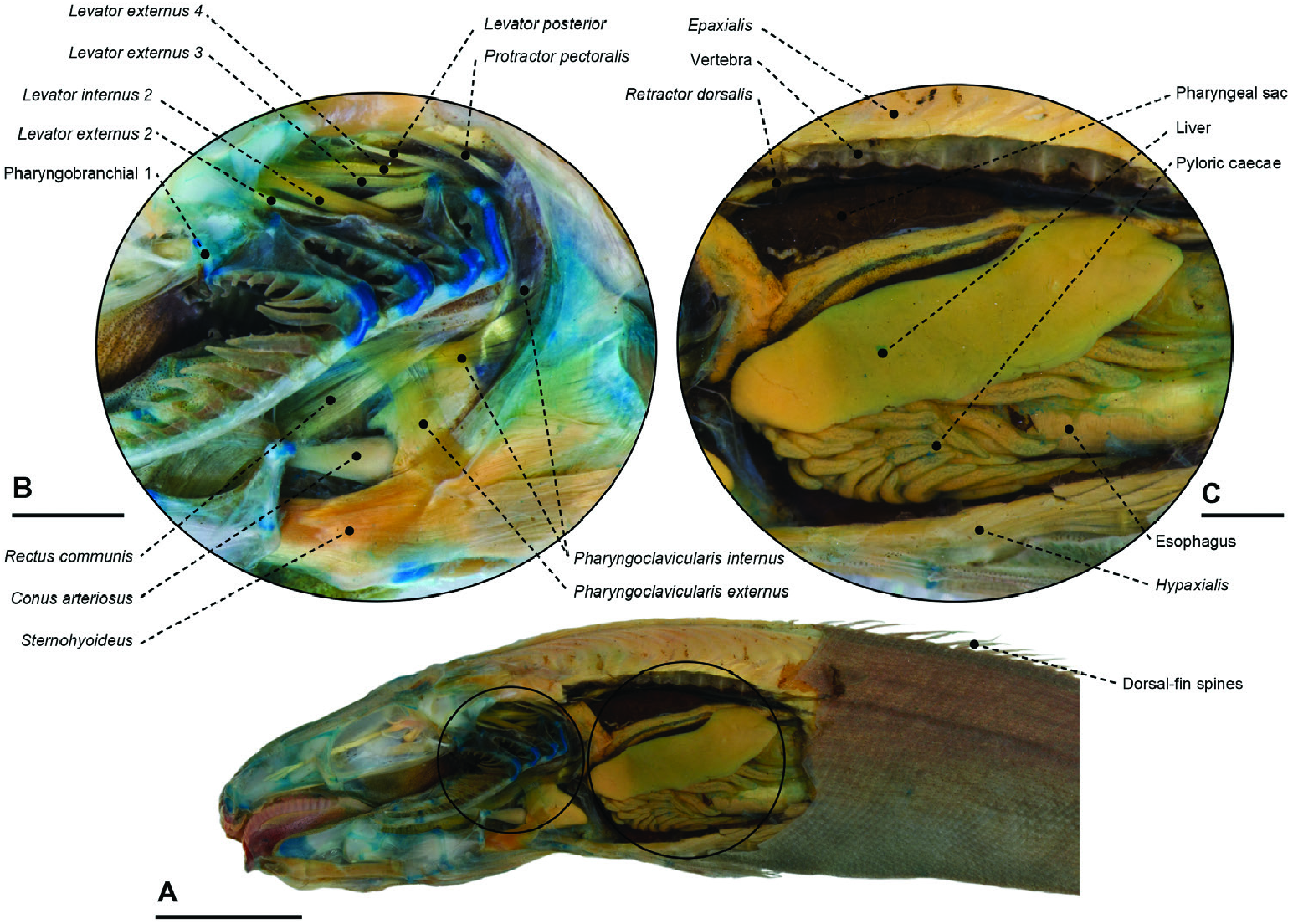
Autapomorphies: Character 0 (11–12> 19): number of dorsal-fin spines increased to 19; character 1 (14> 11): number of dorsal-fin soft rays decreased to 14; character 3 (17–19> 14): number of branched pectoral-fin rays decreased to 14; character 5 (3> 1): number of anal-fin spines decreased to one; character 6 (15> 11): number of anal-fin soft rays decreased to 11; character 9 (33–36> 53): number of vertebrae increased to 53; character 10 (9> 11): number of dorsal procurrent rays increased to 11; character 13 (36.3–36.8%> 41.2%): predorsal length increased to 41.2%; character 14 (60.1–60.6%> 34.8%): dorsal-fin base length decreased to 34.8%; character 15 (24.9– 26%> 13.6%): pectoral-fin length decreased to 13.6%; character 16 (36.1–36.8%> 30.7%): prepelvic length decreased to 30.7%; character 18 (31.3–37.4%> 10.7%): anal-fin base length decreased to 10.7%; character 23 (0> 1): sclerotic bone absent; character 30 (1> 0): medial bony suture between anterior and posterior ceratohyals absent; character 38 (1> 0): interarcual cartilage absent; character 41 (1> 0): vomerine teeth present; character 42 (1> 0): palatine teeth present; character 45 (1> 0): basihyal/hypobranchial/ basibranchial teeth present; character 84 (0> 1): supraneurals absent; character 92 (0> 1): adductor mandibulae pars malaris reaching the dorsolateral edge of the preopercle; character 98 (1> 0): adductor mandibulae partes rictalis and stegalis completely separated; character 100 (0> 1): adductor mandibulae segmentum mandibularis notched anteriorly; character101(1>0):adductormandibulaeparscoronalis not extending past the posterodorsal border of the lower jaw; character 102 (1> 0): adductor mandibulae pars mentalis not differentiated into prementalis and postmentalis sections; character 108 (1> 0): RLA-OP medial to the levator operculi; character 110 (1> 0): dilatator operculi and levator arcus palatini separated only in part; character 131 (0> 1): rectus ventralis I present; character 133 (1> 0): pharyngoclavicularis externus lateral to sternohyoideus; character 135 (0> 1): pharyngoclavicularis internus subdivided; character 139 (1> 0): pharyngoclavicularis internus anteriorly positioned in relationship to pharyngoclavicularis externus; character 146 (1> 0): anterior border of the anterolateral section of the epaxialis not reaching the vertical through the middle of the orbit; character 149 (1> 0): anterior border of the anteromedial section of the epaxialis not reaching the vertical line through the middle of the orbit; character 182 (1> 0): intradermal canal plexus absent; character 199 (1> 2): scales spinoid; character 203 (1> 0): fleshy groove at the ventral profile of the body absent.
Support: –.
Remarks: The family Tetragonuridae comprises one genus and three valid species, Tetragonurus atlanticus Lowe, 1839 , Tetragonurus pacificus Abe, 1953 and Tetragonurus cuvieri ( Haedrich & Horn, 1972; Fricke et al., 2020). The family groups the most modified species of Stromateiformes , as evidenced by its long branch containing 35 autapomorphies ( Fig. 70 View Figure 70 ). These include numerous features unique among stromateiforms, such as the highest number of dorsalfin spines (character 1), the lowest number of analfin spines (character 8), small dorsal- (character 17) and anal-fin base lengths (character 21), absence of supraneurals (character 101), absence of an intradermal canal system on the body (character 187) and presence of spinoid scales (character 206). These fishes additionally exhibit unique characteristics such as a highly modified lower jaw, equipped with teeth that resemble the edge of a saw ( Fig. 48 View Figure 48 ), a poorly ossified cranial skeleton ( Fig. 53 View Figure 53 ) and an imbricate scale pattern along the body ( Fig. 71A View Figure 71 ) that, according to some authors, forms a water-filled channel system analogous to the intradermal canal plexus present on the remaining stromateiforms ( Bone & Brook, 1973). Nevertheless, these characteristics were not included in the present analysis owing to their autapomorphic occurrence in our representative of Tetragonuridae ( Tetragonurus cuvieri ). Tetragonurids also differ from other stromateoids in their primary association with gelatinous invertebrates. Juveniles and young adults of Tetragonurus are always observed associated with salps and pyrosomes, whereas most stromateiforms are symbiotic with jellyfish and ctenophores. According to the behavioural study by Janssen & Harbison (1981), juvenile tetragonurids spend most of their life confined inside salps and pyrosomes, feeding on their guts and gonads. This highly specific symbiotic association with tunicates might be correlated with some of their aberrant morphological modifications, such as the specialized jaw and teeth ( Fig. 48 View Figure 48 ) and the elongate and shallow body ( Fig. 71A View Figure 71 ). The symbiosis of Tetragonurus with tunicates, rather than jellyfishes, might also explain the absence of an intradermal canal plexus on their bodies, because one of the possible functions of this system is to protect against cnidarian stings.
Given the number of morphological modifications present in tetragonurids, it is not surprising that the family has been hypothesized to belong to different lineages of stromateiforms. A Tetragonuridae + Ariommatidae sister-group relationship, as supported herein (and by Horn, 1984), is based mostly on shared pharyngeal-sac specializations (see Remarks on node 92). Alternative Tetragonuridae affinities include a sister-group relationship with stromateids ( sensu Doiuchi et al., 2004; Doiuchi & Nakabo, 2006). Morphologically, this grouping was based on three shared features (anterior and posterior ceratohyals medially separated by a strip of cartilage, pharyngobranchial 3 articulation site with epibranchial 3 shifted anteriorly, and epurals reduced to two; Doiuchi et al., 2004: characters 13, 16 and 25, respectively). Molecular analyses with broader sampling of outgroup taxa offer more disparate hypotheses for the tetragonurid interrelationships. In the studies by Miya et al. (2013) and Friedman et al. (2019), Tetragonuridae is sister to the nonstromateiform Chiasmodontidae , and in the study by Campbell et al. (2018) it is grouped with Amarsipidae in a clade that is closer to scombrids ( Scombriformes ) than to the remaining stromateiforms.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |