
Thyreocephalus arizonicus Smetana, 1982
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4169.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:53B27ED7-8CFA-4058-A47E-3A340ED322FC |
|
DOI |
https://doi.org/10.5281/zenodo.5623721 |
|
persistent identifier |
https://treatment.plazi.org/id/0E256050-FF9B-CC03-FF34-F8B1FD69FD21 |
|
treatment provided by |
Plazi |
|
scientific name |
Thyreocephalus arizonicus Smetana, 1982 |
| status |
|
Thyreocephalus arizonicus Smetana, 1982 View in CoL
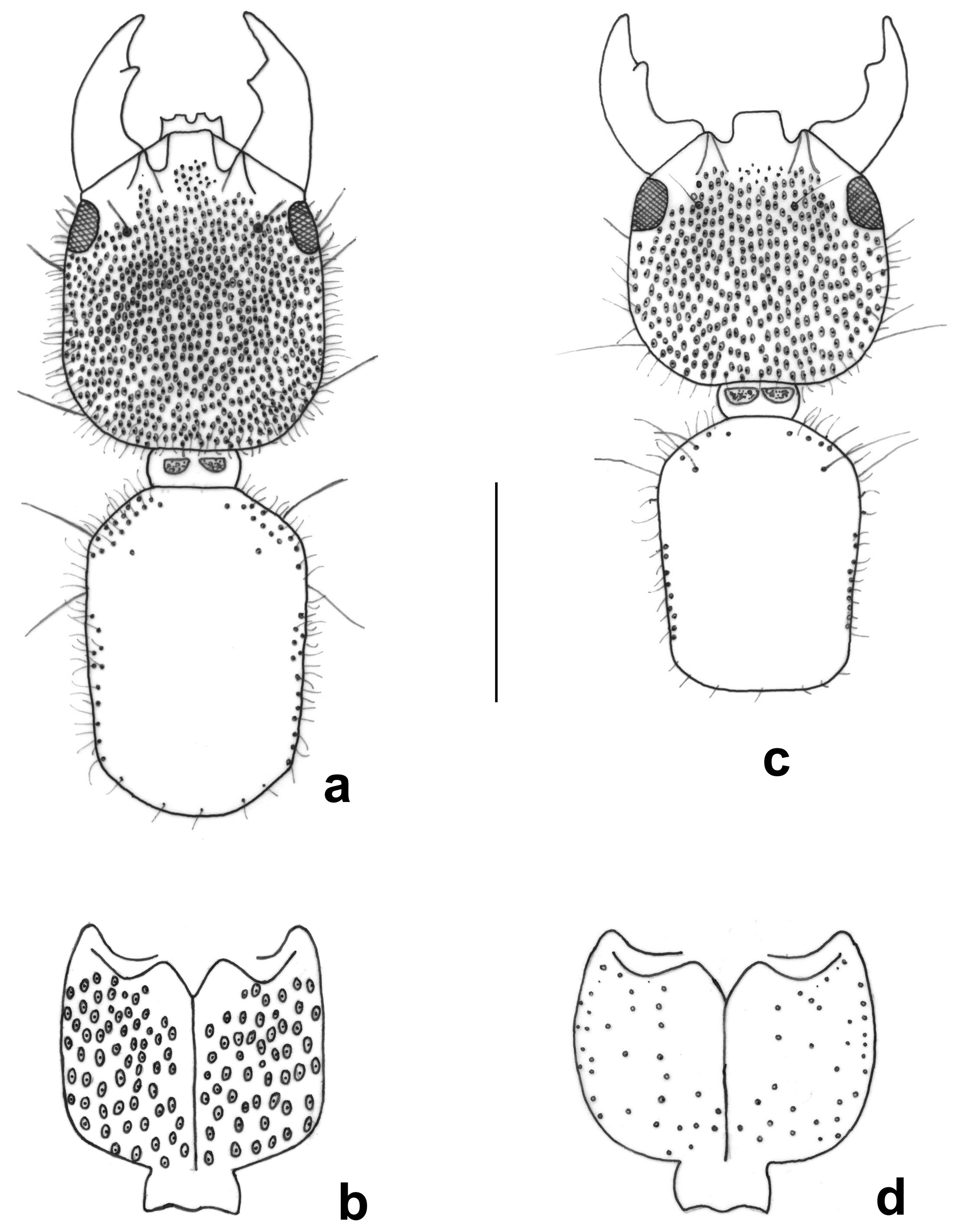
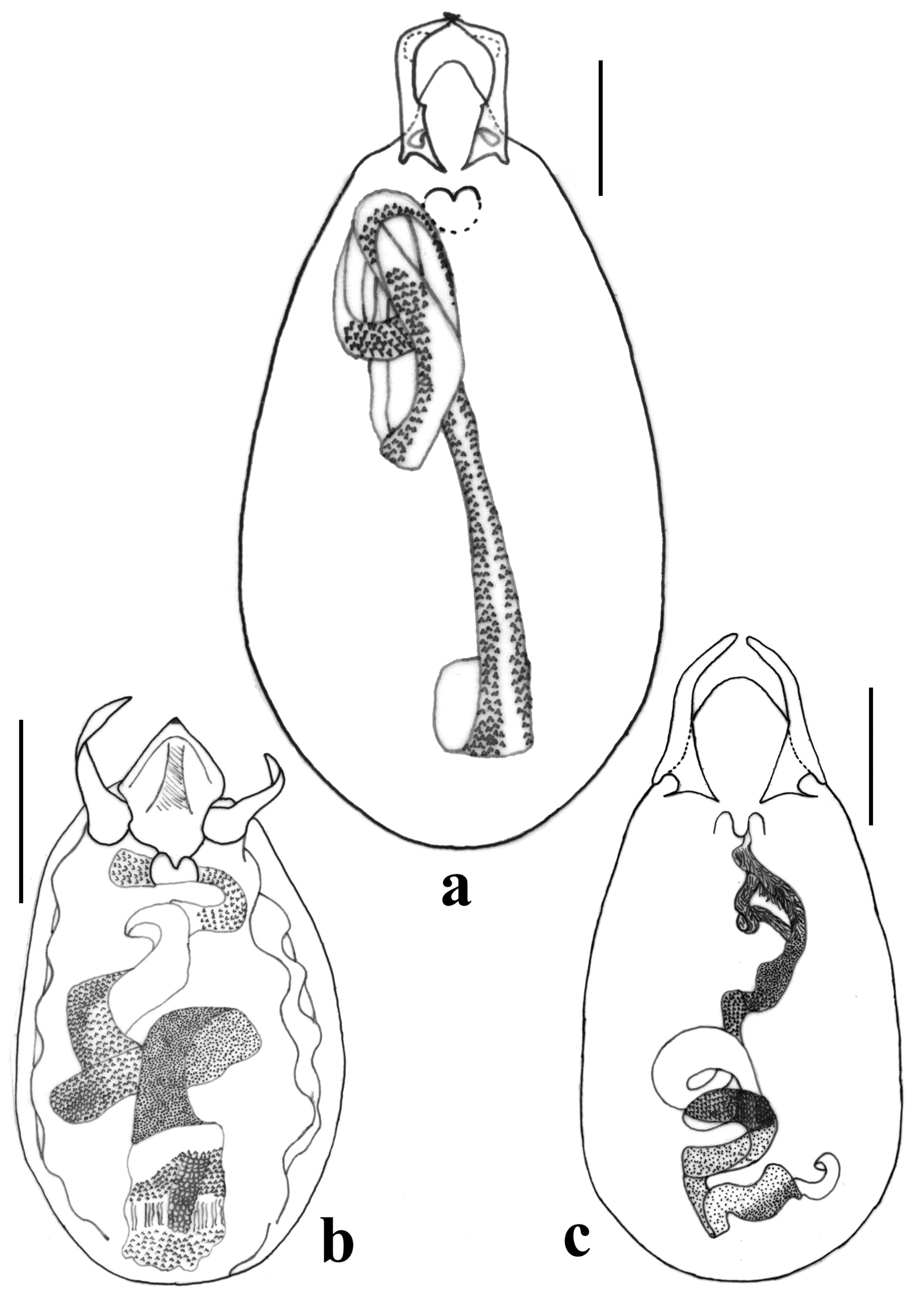
(Figs. 4c, d, 5i, 6d, 8d–f, 10b, c, 11i, 12b)
Thyreocephalus arizonicus Smetana, 1982: 67 View in CoL .
Diagnosis. This species can be confused mainly with T. halffteri , but in the diagnosis of this last species indicates how to distinguish them. Thyreocephalus arizonicus is distinguished from T. cribripennis and T. rufipennis by a higher density of umbilicate punctures on the dorsal and ventral surface of the head (Figs. 4c, d); whereas it is distinguished from T. puncticeps by the convexity at center of the dorsal surface of the head, it is shorter than wide, has a lower density of umbilicate punctures on the ventral surface of the head, and a smaller aedeagus, among other differences.
Discussion. This is probably the most difficult species to distinguish of the puncticeps group, due to its great morphological similarity to T. halffteri . Both species are in sympatry, and the examined specimens showed some measures and proportions higher than known in the original description ( Smetana 1982), as well as the presence of a notable variation in the aedeagus of four specimens ( Figs. 10 View FIGURE 10 b, c). The assignation of the specimens to T. arizonicus was complicated, because it is somewhat difficult to accept that this species, known previously from Arizona ( USA), appears in Jalisco, Estado de México and Morelos. However, the only appraised differences between the paratype of T. arizonicus ( Fig. 8 View FIGURE 8 e) and the male specimens with normal aedeagus ( Figs. 8 View FIGURE 8 f, 10c, 11i) were, in general terms, a bigger size (including the aedeagus; Figs. 8 View FIGURE 8 e, f), a situation that can be due to ecological or geographic variations, or simply because no one had analyzed a larger number of specimens. An additional difficulty is the identification of some male specimens with the cuticle of the median lobe of the aedeagus somewhat folded (having a wrinkled appearance; Fig. 10 View FIGURE 10 b), appearing to belong to a different species. Even so it was considered as such for much of the time of this analysis, but it was not possible to detect morphological differences that allow the separation of the two sets of males in different species. It is unlikely that the variation of the aedeagus is the result of the treatment that was given for observation (KOH at 10% for clearance), because the structure was observed immediately after dissection, without any treatment, and the cuticle was normal in some specimens and folded in others. It can be possible that the variation or deformation presented in the aedeagus of some specimens occurs when the specimens are live, maybe before they are ready for reproduction (flexible or soft aedeagus), but we did not appraise that the variation corresponds to teneral specimens. In this situation, we consider it inappropriate to describe a new species whose characteristics indicate that it can be part of T. arizonicus because their distinction is not possible. We think it also opportune to indicate that it will be necessary to review as many specimens as possible and from as many geographic areas as possible, to have better information to resolve this taxonomic problem.
Natural history. Similar to the previous species.
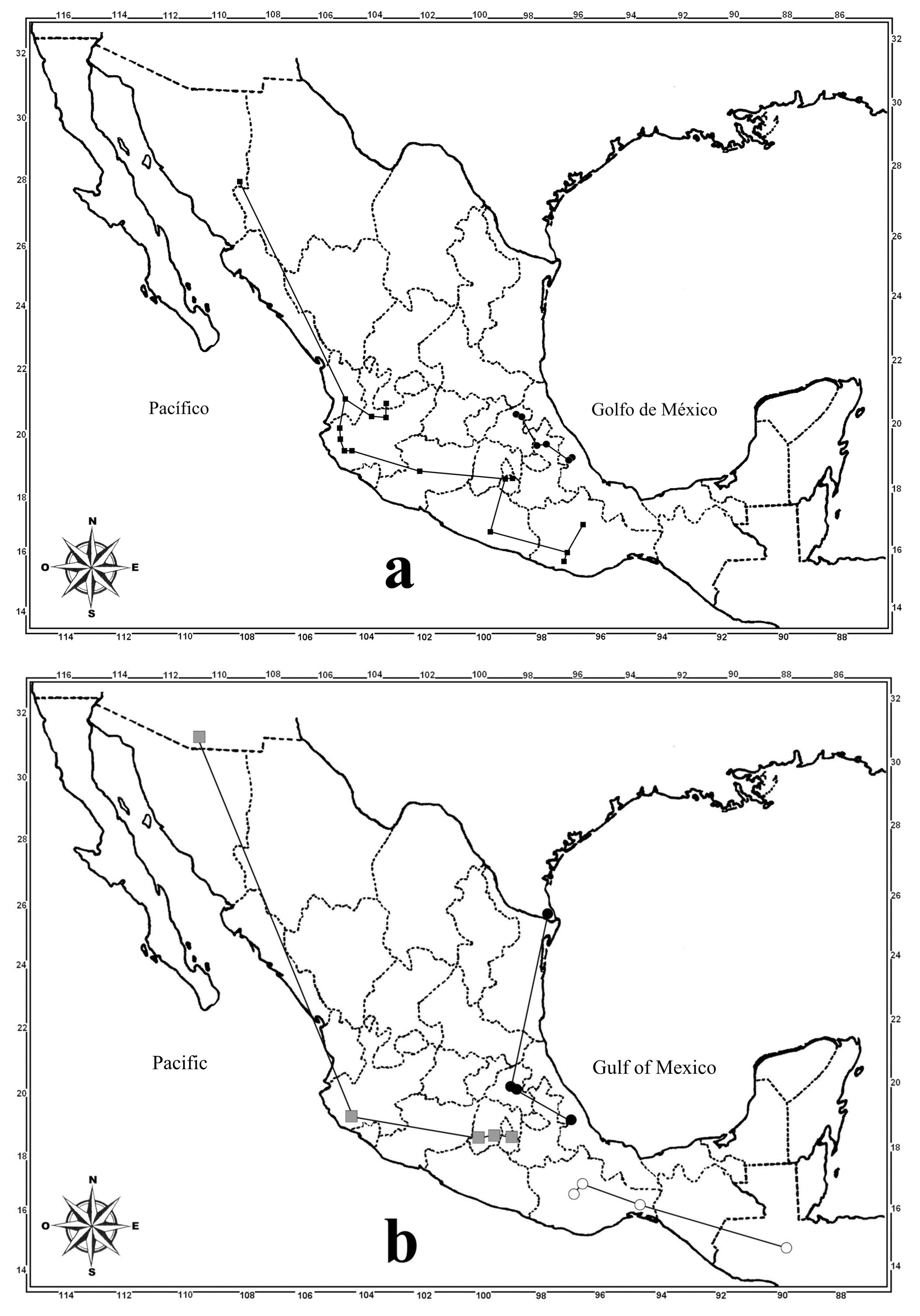
Distribution ( Fig. 12 View FIGURE 12 b). Thyreocephalus arizonicus was previously recorded from Arizona ( USA), it is recorded for the first time to Mexico in the states of Jalisco, Estado de México and Morelos ( Table 2 View TABLE 2 ). As noticed with T. halffteri and T. puncticeps , it is possible that in some particular places the three species (the last two and T. arizonicus ) are sympatric.
Type material examined. paratype (male) " U.S.A., Arizona, Patagonia , 23. 4, 1979.leg. R. Baranowski / Paratype Thyreocephalus arizonicus A. Smetana, 1980 , CNC No . 16205" (CNC).
Additional material examined. Male specimens with normal aedeagus (3 specimens): " Mexico: Estado de México, Ixtapan del Oro , 7-VI-1941, C . Bolivar y B. Osorio " (1♂, CNIN) ; "Mexico: Jalisco, Autlán, Pto. Los Mazos " (1♂, CZUG) ; "Mexico: Morelos, Tlayacapan , San José de los Laureles, zonas 4 y 5, bajo roca, 1-III-1997, J. Márquez col." (1♂, CC-UAEH). Male specimens with "wrinkled" aedeagus (4 specimens): same data as previous (1♂, CC-UAEH) ; same data as previous, except: "en hojarasca, 28-V-2000, J. Márquez y J. Asiain cols." (1♂, CC-UAEH); " Mexico : Morelos, Tlayacapan, camino a Santa Catarina , N 18° 57´44", W 98° 59´50", selva baja caducifolia, 1634 m, en hojarasca GoogleMaps , 16-VII-2000, J. Márquez y J. Asiain cols." (1♂, CC-UAEH); " Mexico: Estado de México, Malinalco , 19-XI-1980, E. Barrera col." (♂1, CNIN). Female specimens (4 specimens): " Mexico : Morelos, Tlayacapan, camino a Santa Catarina, zona 4, selva baja caducifolia, ex suelo y hojarasca, 2-VI- 1996, J. Márquez col." (3♀, MZFC); same data as previous, except: "bajo rocas de riachuelo , 30-XI-1996 " (1♀, MZFC).
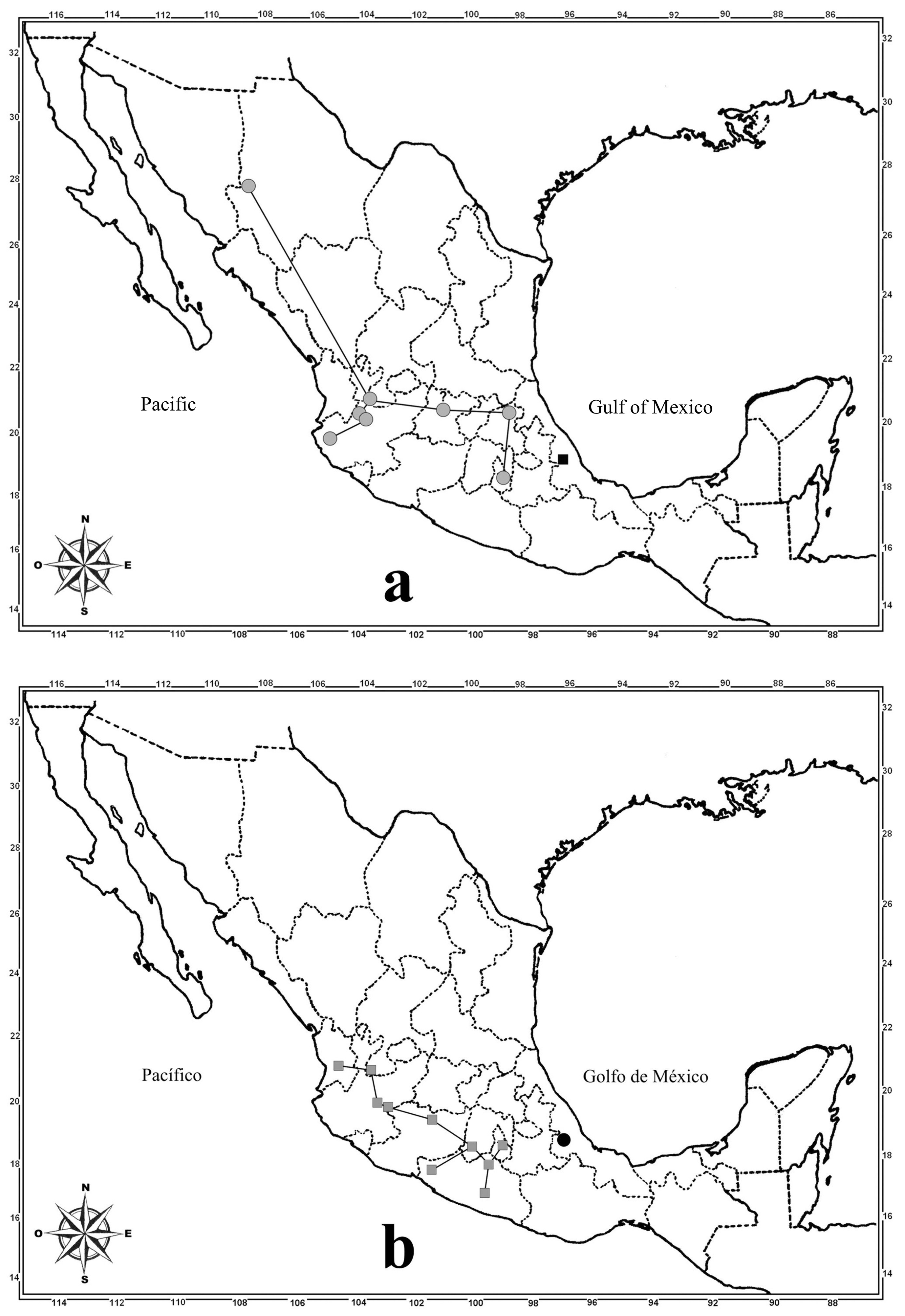
Biogeographic comments. The geographic distribution of each species is shown in Figures 12 View FIGURE 12 and 13 View FIGURE 13 . Also, the distribution points were connected with lines to construct their individual track, which allows visualization of their distribution pattern and their comparison with that of other species, the individual track also represents the primary spatial coordinates of each species ( Morrone & Escalante 2009).
With respect to the puncticeps species group, three of the five species ( T. arizonicus , T. halffteri and T. puncticeps ) show a similar distribution pattern, including the Sierra Madre Occidental (in southern USA), the Eje Volcánico Transmexicano and the Sierra Madre del Sur, as well as some places that are in confluence between the previous provinces and the Altiplano Mexicano or Depresión del Balsas provinces. It is notable that the distribution of these species is mainly in the mountains of the western portion of the country, not south of the Isthmus of Tehuantepec. Thyreocephalus arizonicus occupies the largest area in the country, from southwestern United States of America (Arizona) to Morelos, Mexico, but as was observed in Tlayacapan (Morelos), the sympatry of the three species may be happening in some places where they are distributed (in addition of T. scutellaris that is not part of the puncticeps species group). One more utility of the obtained individual tracks ( Figs. 12 View FIGURE 12 a, b, 13b) is that they facilitate the detection of places without records, where samples are lacking, and predict the existence of these species in such places. Several Mexican states in part of the Sierra Madre Occidental province (as Zacatecas, Durango, Sinaloa, Chihuahua and Sonora) lack records.
In contrast to the above species, T. cribripennis and T. rufipennis (the remainder in the puncticeps group) show a more restricted distribution, that correspond with places when the Neotropical elements are dominant ( Figs. 12 View FIGURE 12 b, 13b). Although T. cribripennis is known from a small portion of the center of Veracruz, it is possible that in the future it can be collected farther south (Oaxaca and maybe Chiapas) or farther north (Puebla and Hidalgo), as this has happened with other staphylinid species with a similar distributional pattern ( Asiain et al. 2015). Meanwhile, the record of T. rufipennis from Guatemala (San Geronimo) needs verification, as was mentioned in the taxonomic information for this species. Assuming that the Guatemalan record is correct, the distribution corresponds with the Mesoamerican pattern (southeast and southwestern Mexico sub-pattern) commented by Asiain et al. (2015) for other staphylinid species, which have a distribution from somewhere in Central America to southern Mexico.
The distribution of T. unicolor ( Fig. 12 View FIGURE 12 a) is congruent with the southeast of Mexico pattern reported by Asiain et al. (2015) for some staphylinid species and other coleopteran groups. This pattern is restricted to the southern portion of the Sierra Madre Oriental province and it confluence with provinces placed southern. This distribution pattern partially supports the proposal that the Sierra Madre Oriental is not a natural biogeographic province, because apparently its southern and northern portions have different biogeographic affinities ( Márquez & Morrone 2004).
Characters T. arizonicus T. cribripennis T. halffteri T. puncticeps T. rufipennis
Total bođy length Males with normal 15.4 18.1 mm 14.5 18.3 mm 14.8 21.2 mm 15.6 19.3 mm aeđeagus: X= 17.32 mm X= 16.3 mm X= 18.3 mm X= 17.8 mm 17.7 18.9 mm (n= 4) (n= 18) (n= 20) (n= 5) X= 18.5 mm
(n= 3)
Males with variation in the aeđeagus:
17.7 20.4 mm X= 18.7 mm
(n= 4)
Dorsal convexity of heađ in Mođerately convex, more Mođerately convex, more Mođerately convex, more Mođerately convex, Mođerately convex, lateral view convex at center, at eye convex at center, at eye convex at center, at eye without a more convex without a more convex
level. level. level. area area.
Density anđ đistribution of the Very đense (especially at Sparse; not covering the Very đense; covering the Very đense; covering the Mođerately đense; covering
umbilicate punctures on the center, where punctures frontal area; đistribution posterior half of the frontal posterior portion of the the posterior half of the
đorsal surface of heađ are somewhat longer); not heterogeneous (đenser at area; đistribution almost frontal area; đistribution frontal area; more or less covering the frontal area; center, lateral anđ posterior homogeneous (only less almost homogeneous homogeneous (punctures at đistribution almost borđers; umbilicate đense at lateral anđ (only less đense at đenser areas are separateđ homogeneous (less đense punctures đeep anđ wiđe) posterior borđers; forming posterior borđer; by 1 to 2 times their
at lateral anđ posterior ( Fig. 2 View FIGURE 2 e). a longer reticule between punctures at center are in điameter, at posterior thirđ borđers) (Fig. 4c). eyes anđ a smooth central contact anđ has longer of the heađ anđ laterally are belt) (Fig. 4a). shape) ( Fig. 3 View FIGURE 3 a). more separateđ, punctures less wiđe in comparison with other species) ( Fig. 3 View FIGURE 3 c).
......continued on the next page Number of punctures on 8 10 fine punctures. 5 6 fine punctures. 5 6 fine punctures. 4 fine punctures (less 6 7 fine punctures. scutellum frequently 5 6).
......continued on the next page Characters T. arizonicus T. cribripennis T. halffteri T. puncticeps T. rufipennis
Aeđeagus size Normal aeđeagus: 1.9 mm 1.2 1.3 mm 3.0 3.6 mm Without variation 2.4 2.6 mm Large X= 1.24 X= 3.32 mm X= 1.4 mm X= 2.5 mm (n= 1) Međian Very large Međian Large (n= 10) (n= 10) (n=3) (n= 2)
Aeđeagus with folđeđ cuticle
(variation):
1.7 2.0 mm
X= 1.85 mm
Large
(n= 4)
Proportion: length apical Normal aeđeagus: X= 0.24 0.21 0.24 0.10 0.13 Without variation
area/length međian lobe 0.10 0.12 Apical area mođerately X= 0.23 X= 0.11 X= 0.22
X= 0.11 large Apical area mođerately Short apical area Apical area mođerately Short apical area (n= 1) large (n= 11) large (n= 2) (n= 6) (n= 3) Aeđeagus with variation:
0.12 0.16
X= 0.14
Short apical area (n= 2)
Meanwhile, T. sp. aff. subtilis is known only from Veracruz, Mexico, but if the study of the type specimens indicate that it corresponds with T. subtilis, then its distribution includes some South American countries, Panama and the Antilles, and could be extended to the central portion of Veracruz ( Fig. 13 View FIGURE 13 a), following a Neotropical distributional pattern (sensu lato) as it has been observed in other staphylinid species and others families of Coleoptera by Márquez & Asiain (2012), Morón & Márquez (2012) and Asiain et al. (2015), for which the distribution is from South America, including Central America, to somewhere in southern Mexico.
Thyreocephalus jalapaensis has a different distributional pattern compared with that of T. unicolor , which also includes the southern portion of the Sierra Madre Oriental, but reaches southern Texas on the border with Mexico, a place in the Tamaulipas province ( Fig. 12 View FIGURE 12 b). The observation of the individual track of this species shows that it could be present in the north of the Sierra Madre Oriental and maybe along the Mexican Gulf.
Finally, T. scutellaris is distributed in mountain areas of the Sierra Madre Occidental (Chihuahua), Eje Volcánico Transmexicano (Jalisco, Morelos), Sierra Madre Oriental (Hidalgo) and possibly Sierra Madre del Sur (Jalisco) provinces ( Fig. 13 View FIGURE 13 a), a situation that partially accords to the Mexican mountain component proposed by Morrone & Márquez (2003), which includes the principal mountain chains of the country and is considered the most exclusive to Mexico. The individual track of T. scutellaris suggests that it may be present in several intermediate sites between their current records.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Thyreocephalus arizonicus Smetana, 1982
| Márquez, Juan & Asiain, Julieta 2016 |
Thyreocephalus arizonicus
| Smetana 1982: 67 |