
Euphysilla pyramidata Kramp, 1955
|
publication ID |
https://doi.org/ 10.1163/18759866-BJA10023 |
|
DOI |
https://doi.org/10.5281/zenodo.8357081 |
|
persistent identifier |
https://treatment.plazi.org/id/12088786-8E23-FF8A-FF91-637CE5C4FB1D |
|
treatment provided by |
Felipe |
|
scientific name |
Euphysilla pyramidata Kramp, 1955 |
| status |
|
Euphysilla pyramidata Kramp, 1955 View in CoL View at ENA
Euphysilla pyramidata Kramp, 1955: 245 View in CoL , pl. 1 fig. 1 View FIGURE 1 , pl. 2 fig. 3 View FIGURE 3 ; Kramp, 1965: 4; Vannucci & Santhakumari, 1969: 40; Schmidt, 1973: 16; Hamond, 1974: 554, figs 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 ; Segura-Puertas, 1984: pl. 2 fig. 2 View FIGURE 2 ; Buecher & Gibbons, 2000: 127; Xu & Huang, 2004: 560, fig. 9 View FIGURE 9 ; Schuchert & Collins, 2021: 265 View Cited Treatment , fig. 17.
Sphaerocoryne sp. Di Camillo et al. 2008: 1591; Maggioni et al. 2017: 737.
Examined material: Sample sp002, Maldives, April 2015, polyps in ethanol. – Sample MA 16001, Maldives, 19/01/2016, polyps in ethanol and formalin. – Sample MA 16007, Maldives, 21/01/2016, polyps in ethanol. – Sample MA 16014, Maldives, 22/01/2016, polyps in ethanol and formalin. – Sample MA 16022, Maldives, 25/01/2016, polyps in ethanol and formalin. – Sample MA 16039, Maldives, 29/01/2016, polyps in ethanol. – Sample MA 16054, Maldives, 09/02/2016, polyps in ethanol and formalin. – Sample MA 0117032, Maldives, 31/01/2017, polyps in ethanol. – Sample MA 0117133, Maldives, 13/02/2017, polyps in ethanol and formalin (MHNG-INVE-0137430). – Sample STE040, Sint Eustatius, 18/06/2015, polyps in ethanol.– Sample BFLA4396 , off Florida, 17/05/2020, medusa in ethanol and in situ photos. – Sample BFLA4397 , off Florida, 17/05/2020, medusa in ethanol and in situ photos. – Sample BFLA4478 , off Florida, 18/06/2020, medusa in ethanol and in situ photos.
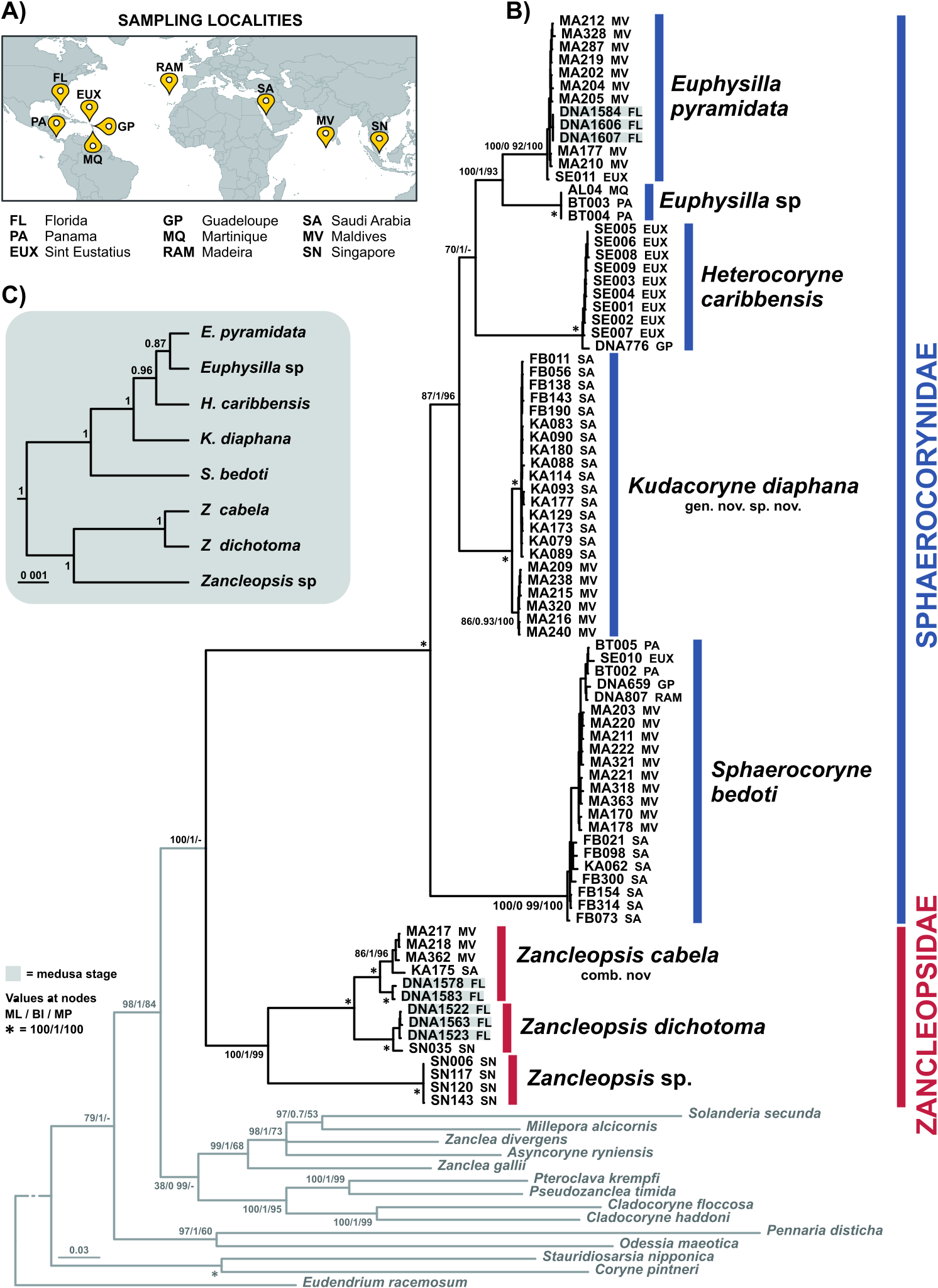
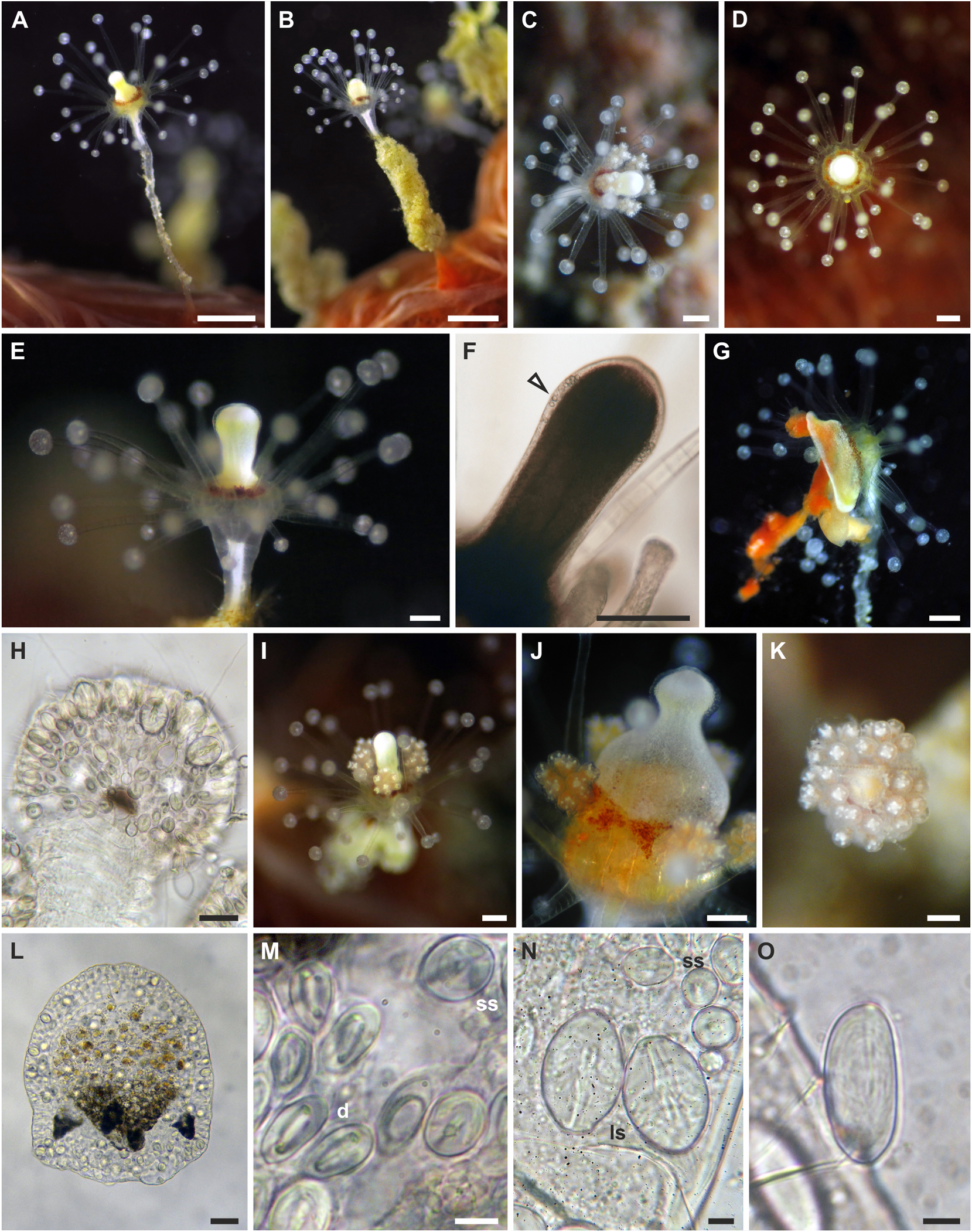
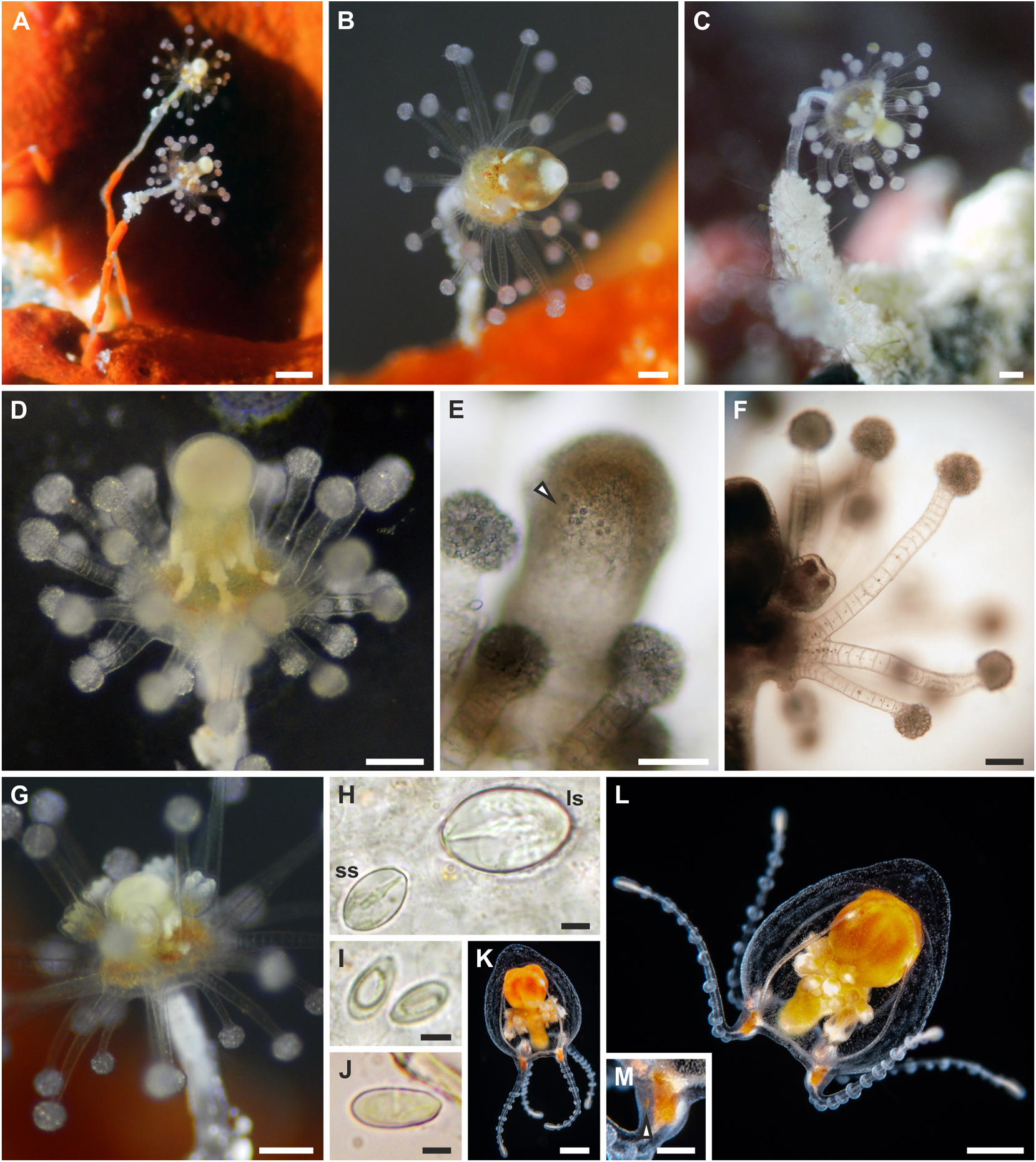
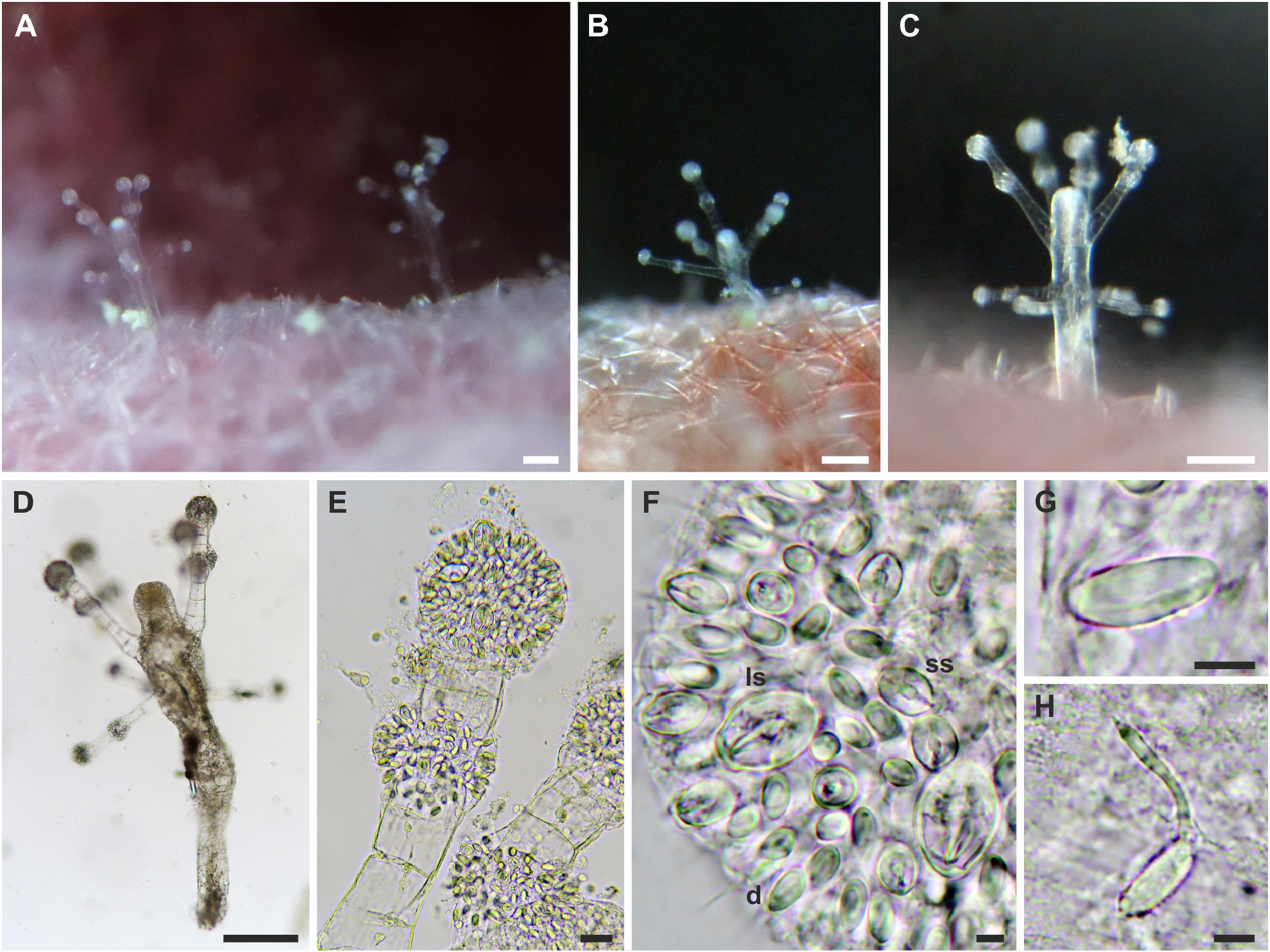
Description: Polyp. Colonies monomorphic, living in association with sponges and occasionally growing on carbonatic rock ( fig. View FIGURE 4 4A-C). Hydrorhiza tubular, branched, covered by perisarc and growing in the sponge host. Pedicels long (up to 5 mm), unbranched, covered by a smooth, thin perisarc ( fig. 4A View FIGURE 4 ), occasionally highly covered by debris ( fig. 4C View FIGURE 4 ). Hydranth pyriform, up to 2.5 mm long, with variable diameter (150–420 Μm) ( figs. 4 View FIGURE 4 A-D), in some cases directed at an angle of 90° to the pedicel ( figs. 4B, C View FIGURE 4 ). Hypostome proboscis-like, contractile, with a band of nematocysts below the mouth ( fig. 4E View FIGURE 4 ). Up to 45 tentacles arranged in 3–5 close whorls in the broadest part of the polyp, and grouped longitudinally ( fig. 4F View FIGURE 4 ). Each tentacle with a terminal, nematocyst-rich capitulum (diameter: 100–140 Μm in the distal whorls; 85–95 Μm in the proximal whorl). Tentacles up to 600 Μm long in the distal whorls, shorter in the proximal whorl. Up to seven medusa buds at the same stage of maturation develop above distal tentacles, singly on blastostyles ( fig. View FIGURE 4 4G). Living hydranths with white mouths, an irregular yellow-orange band below the hypostome, at the level of the gastric cavity, and white, clearly visible, mesenteries ( figs. View FIGURE 4 4A-D). Small and large stenoteles ( fig. 4H View FIGURE 4 ) and desmonemes ( fig. 4I View FIGURE 4 ) occurring simultaneously and concentrated in the capitula, and rarely also scattered in the hydrorhiza and in the hydranth; heteronemes rarely found in the hydrocaulus ( fig. 4J View FIGURE 4 ); small stenoteles in a band around the hypostome.
Downloaded from Brill.com 10/07/2022 07:01:58 PM via free access
Polyp cnidome. i) Desmonemes (undischarged: 10 × 4–5 Μm; discharged capsule: 8 × 5 Μm). ii) Heteronemes (undischarged: 16 × 7). iii) Large stenoteles (undischarged: 22–24 × 12–15 Μm; discharged capsule: 19–21 × 12–13 Μm). iv) Small stenoteles (undischarged: 11–14 × 8–10 Μm; discharged capsule: 10 × 7–8 Μm).
Downloaded from Brill.com 10/07/2022 07:01:58 PM via free access
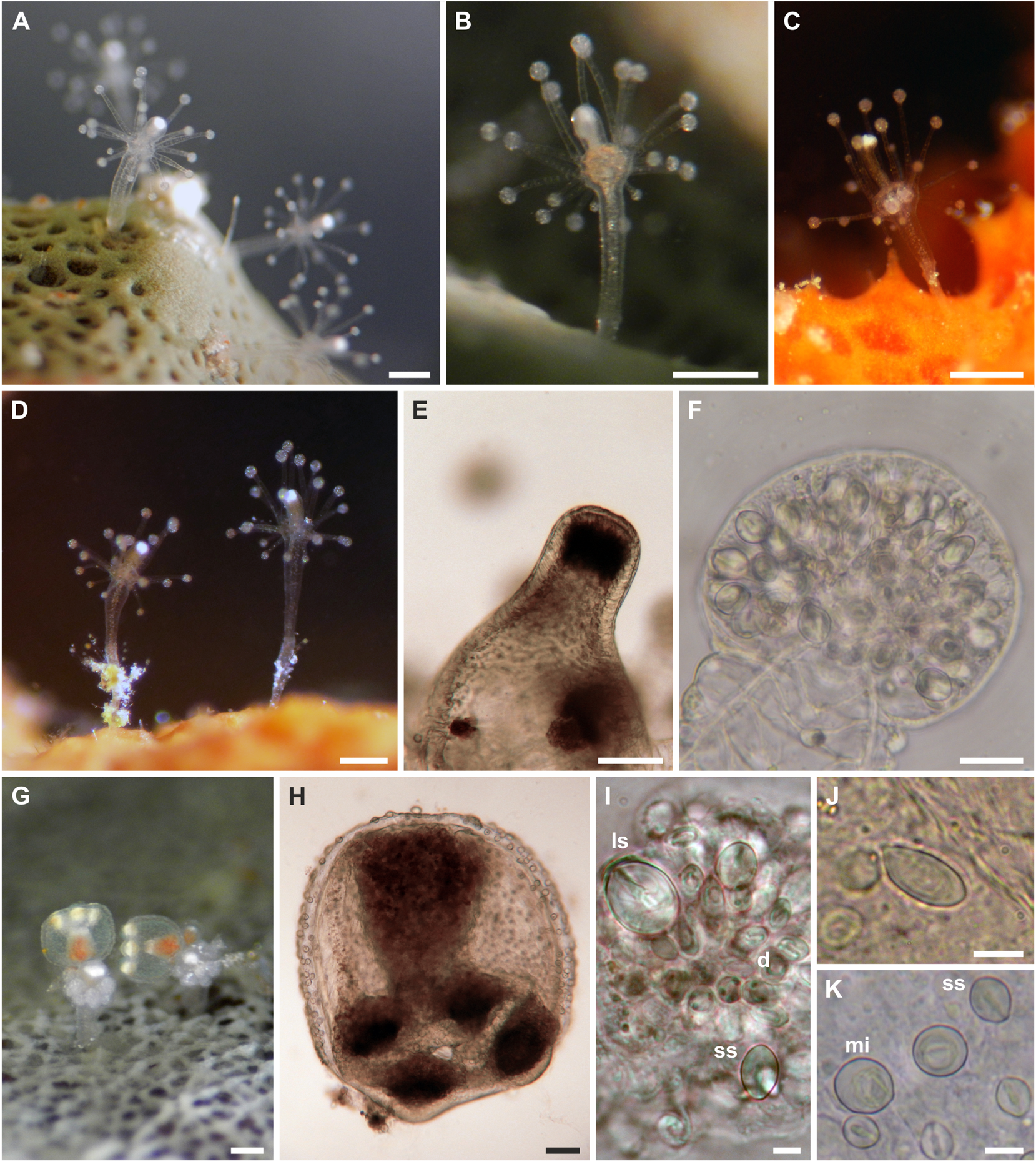
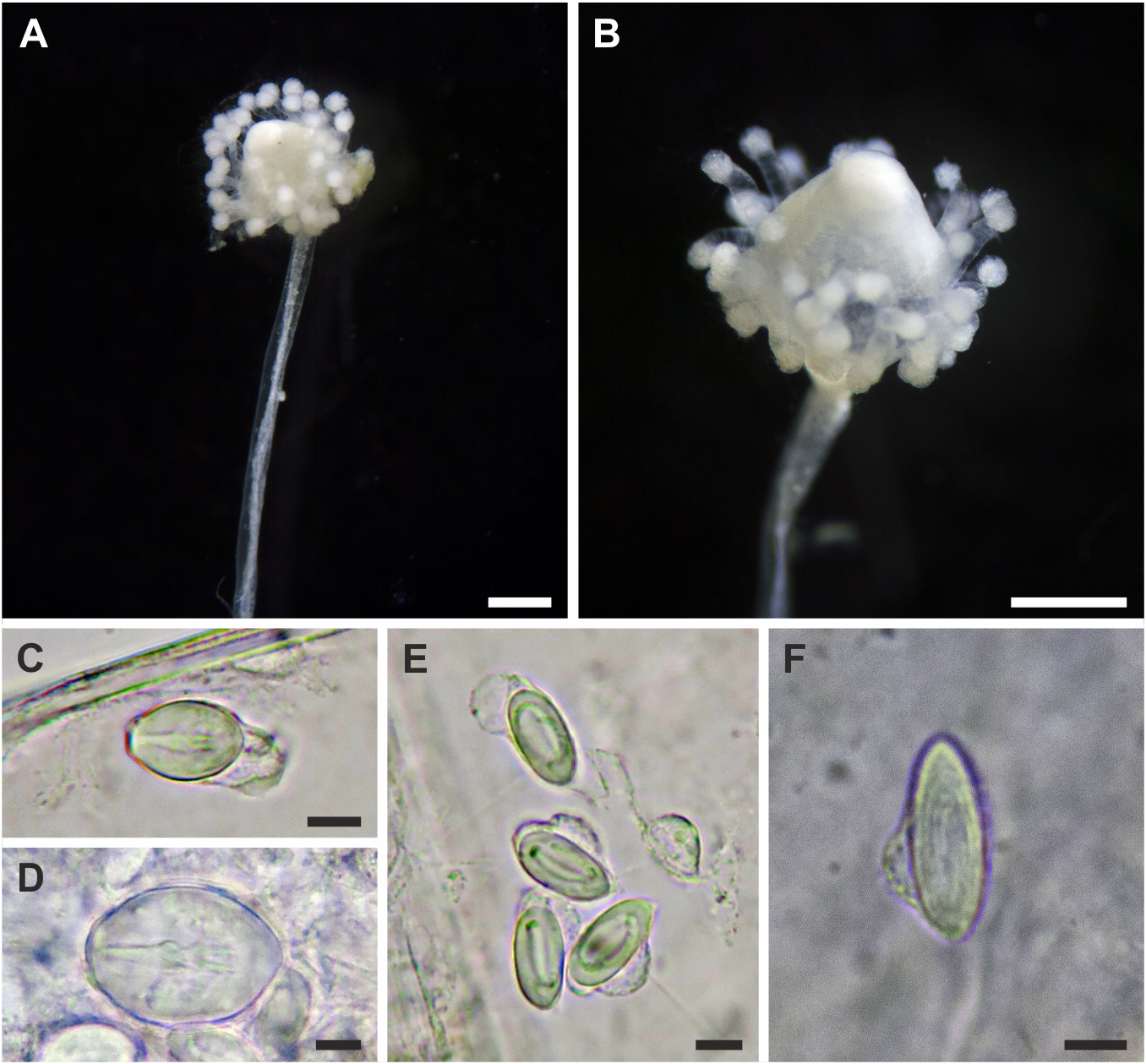
Adult medusa (from Schuchert & Collins, 2021). Umbrella 2–4 mm in height, oviform, with apical thickening of very variable height, lateral walls thin, exumbrella with scattered nematocysts ( fig. 4K, L View FIGURE 4 ). Above manubrium an apical chamber, without apparent connection to gastric cavity, size variable depending on size of apical process. Manubrium an inverted cone, as long as bell cavity, base broad and cruciform in life, more square-shaped when preserved, upper part of manubrium cylindrical, narrowing to tubular lower part of manubrium ending in small, circular mouth. All observed medusae were budding medusae, buds in groups on all four perradial sides of the manubrium in about the middle of the manubrium. Gonad-like opaque tissue layer covers manubrium above buds. Colour of manubrium intensively yellow-orange. Radial canals connected to manubrium by apparent short mesenteries (giving cruciform manubrium base), thin; circular canal more rectangular than circular. Four tentacles, contracted about half the length of the bell height, each with 8–12 crescent-shaped, clasping nematocyst pads, all in one row on adaxial side, terminal button ovoid, as wide as rest of tentacle. Four tentacle bulbs relatively small, orange-yellow, with inconspicuous orange-yellow dots on abaxial side of tentacle bulbs resembling reduced ocelli ( fig 4M View FIGURE 4 ).
Adult medusa cnidome (preserved material). i) Desmonemes (4 × 9 µm). ii) Small stenoteles (7 × 10 µm). iii) Large stenoteles (12 × 13 µm). iv) Spherical microbasic euryteles with barbed filament, shaft appears without barbs (7 × 10 µm).
Distribution: Atlantic Ocean and Caribbean Sea ( Benin, Florida; Sint Eustatius), Red Sea and Indo-West Pacific (Northern Red Sea; Australia, India, Indonesia, Madagascar, Maldives, South Africa, Taiwan Strait), and East Pacific. The records from Papua New Guinea may refer to another species ( Schuchert & Collins, 2021).
Remarks: The polyp stage of E. pyramidata is herein described for the first time, and largely resembles Sphaerocoryne and Kudacoryne polyps. However, differences are found in the colouration of the hydranth, tentacle organisation, lack of inclusions in the capitula, and organisation and number of medusa buds. It is possible that polyps of this species were assigned to the genus Sphaerocoryne in previous works, as for instance Sphaerocoryne sp. from Indonesia by Di Camillo et al. (2008) (CG Di Camillo, pers. comm.). The medusa shows strong similarities to Sphaerocoryne medusae as well, but is distinguished by the absence or reduction of ocelli and the common presence of medusa buds on the manubrium. Schuchert & Collins (2021) initially detected the genetic similarities in the 16S rRNA region between a Sphaerocoryne polyp from Maldives ( Maggioni et al., 2017) and E. pyramidata medusae from Florida, here confirmed on the basis of multiple DNA regions and phylogenetic analyses. They also suggested that E. pyramidata in the sense of Bouillon (1978) may be a species complex due to morphological differences, but further sampling is still needed to address this issue.

| PM |
Pratt Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Capitata |
|
Family |
|
|
Genus |
Euphysilla pyramidata Kramp, 1955
| Maggioni, Davide, Schuchert, Peter, Arrigoni, Roberto, Hoeksema, Bert W., Huang, Danwei, Strona, Giovanni, Seveso, Davide, Berumen, Michael L., Montalbetti, Enrico, Collins, Richard, Galli, Paolo & Montano, Simone 2021 |
Sphaerocoryne sp.
| Maggioni, D. & Galli, P. & Berumen, M. L. & Arrigoni, R. & Seveso, D. & Montano, S. 2017: 737 |
| Di Camillo, C. G. & Bavestrello, G. & Valisano, L. & Puce, S. 2008: 1591 |
Euphysilla pyramidata
| Schuchert, P. & Collins, R. 2021: 265 |
| Xu, Z. Z. & Huang, J. Q. 2004: 560 |
| Buecher, E. & Gibbons, M. J. 2000: 127 |
| Hamond, R. 1974: 554 |
| Schmidt, H. E. 1973: 16 |
| Vannucci, M. & Santhakumari, V. 1969: 40 |
| Kramp, P. L. 1965: 4 |
| Kramp, P. L. 1955: 245 |