
Pytinicarpa tonitrui Lannuzel, Gâteblé & M.Pignal, 2022
|
publication ID |
https://doi.org/10.11646/phytotaxa.574.2.1 |
|
DOI |
https://doi.org/10.5281/zenodo.7383480 |
|
persistent identifier |
https://treatment.plazi.org/id/1258636B-FFD4-4439-E88E-FF7AFE67A8B3 |
|
treatment provided by |
Plazi |
|
scientific name |
Pytinicarpa tonitrui Lannuzel, Gâteblé & M.Pignal |
| status |
sp. nov. |
Pytinicarpa tonitrui Lannuzel, Gâteblé & M.Pignal View in CoL , sp. nov. ( Figs. 8 View FIGURE 8 , 9 View FIGURE 9 )
Diagnosis:— Pytinicarpae sarasinii (Däniker) G.L.Nesom similis, magnitudine formaque, sed tubo corolla florum ligulatorum omnino glabro et tubulosis bisexualibus floribus carentibus, praecipue differt. Similis Pytinicarpae kaalaensi Lannuzel, Gâteblé & M.Pignal, sed habitu rosulanti multo longioribus folisque (2–5 vs 6–11 cm) praecipue differt.
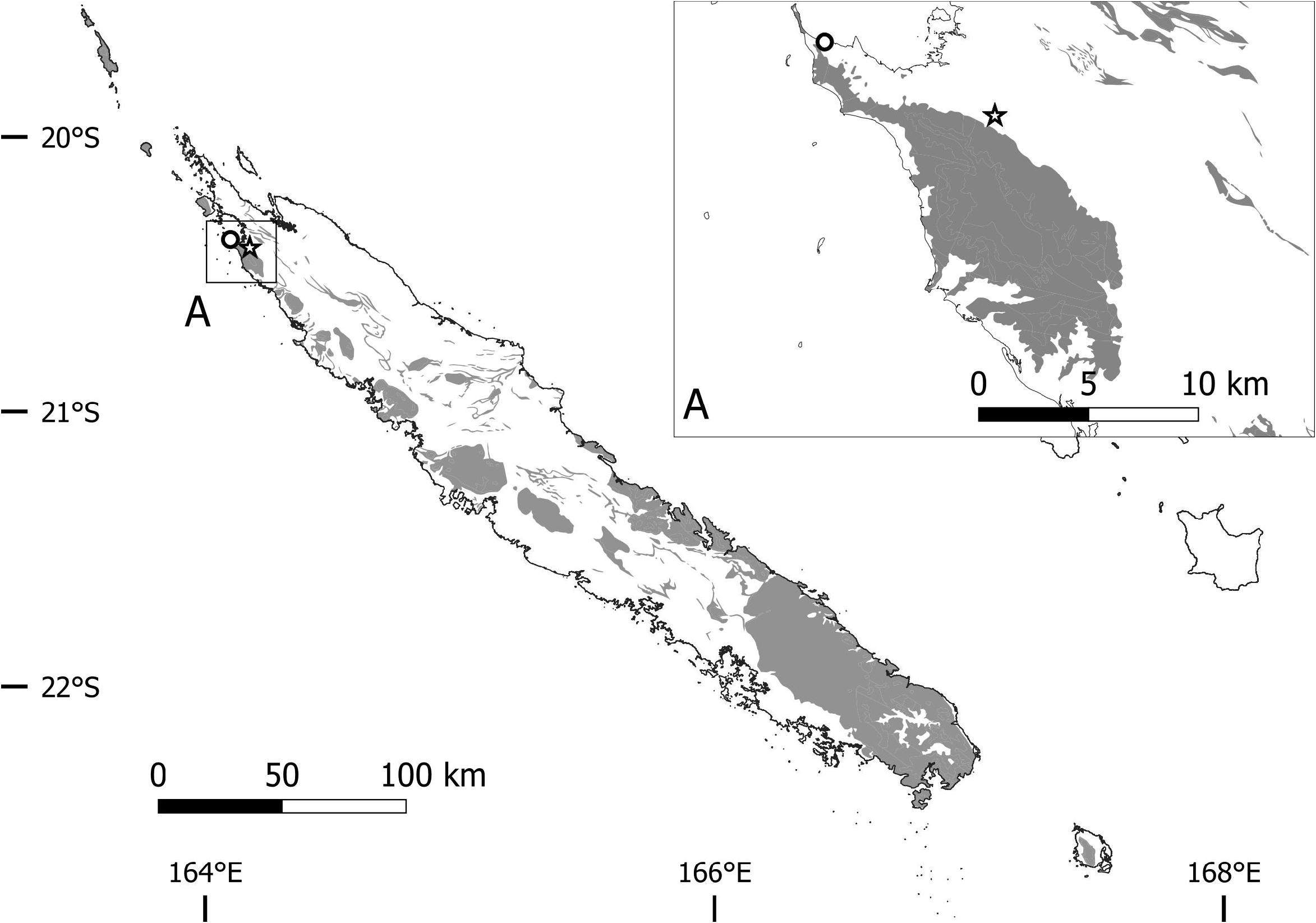
Type :— NEW CALEDONIA. North Prov. : Néhoué, Babouillat, 04 February 1968, H. S . MacKee 18336 ( Holotype: P barcode P03276809 !) .
Perennial rhizomatous herbs; roots fibrous; above ground stem absent (leaves in basal rosette). Leaves 5–15, linear to narrowly oblong, (44–) 79–130 (–162) mm long × (2–)3–5(–6) mm wide (22–27× longer than wide), lamina attenuate to the base, petiole indistinct; leaf apex obtuse with a terminal tooth; margins entire or dentate, with 1–3 teeth per side, each tooth ca. 0.5 mm long when present; upper leaf surface dark greyish-green on dry material; lower leaf surface pale greyish-green on dry material, both surfaces glabrous, but trichomes ( 1–2 mm) sometimes present at leaf base; leaf margins and surfaces glabrous to hairy with trichomes 1–1.5 mm; secondary veins brochidodromous, obscure, sometimes visible on abaxial surface on dry material. Scapes generally 1–3 per tuft, 8–23 cm long, (0.3–) 0.5–0.9 (–1.4) mm diameter; bracts 2–3, 1.8–6.4 × 0.2–0.5 mm with trichomes 0.4–0.8 mm long; 0–1 trichomes per mm² at mid-point of scape, glabrous towards apex; Capitula ca. 3 mm long, (3.4–) 4.8–7.2 (–9) mm diameter; phyllaries ca. 30 in 2–3 rows, ovate to ovate-lanceolate, apex obtuse to acute, glabrous, margins glabrous to ciliate on distal half, outer phyllaries 3 × 1 mm. Receptacle sharply conical at anthesis, 2.5–3 mm diameter and 1.5–2 mm high. Ray florets female 18–22 in 1 row; tube 0.5–1 mm long, ca. 0.2 mm wide, glabrous; style branches ca. 1 mm long; ligules 5–8.2 × 0.7–1.7 mm, with longitudinal veins obscure, glabrous, apex obtuse, ovary 1.1–2.4 × ca. 0.4 mm. Disc florets male tubular, ca. 1.3 mm long, outer surface glabrous; corolla lobes 5, deltate, 0.6–0.8 × 0.7–0.8 mm; stamens, 5, anthers yellow, linear, ca. 0.8 mm; style branches ca. 2 mm long; sterile ovary ca. 1.2 mm long, pappus scales absent. Cypselae oval in cross section, lacking beak, obfusiform, 3.2 × 0.8 mm uniformly brown at maturity; surfaces with 2–3 longitudinal ribs on each side.
Distribution and habitat:—The species is only known from Néhoué–Babouillat (Neué) on the north base of the ultramafic Tiebaghi massif or Cap Tonnerre. Despite several days of fieldwork at different periods of the years in search of this rare species, we were unable to find it. From MacKee 18336 and specimens collected immediately before and after, the ecology of the Babouillat population could be inferred as an open scrubland (maquis minier) at low elevation, most probably on serpentinic alluvium.
Phenology:—From the most recent herbarium specimen with only one mentioning a precise date, the species flowers at least in early February.
Etymology:—This species is named after Cap Tonnerre (“Thunder Cape” in English), a locally accepted name for the peninsula where the locality Néhoué-Babouillat is located, and where every known specimen were most probably collected.
Conservation status:—The Néhoué–Babouillat locality and population where the type of Pytinicarpa tonitrui was collected could be considered as extinct. MacKee label, in 1968, mentions that “the species seems to be rare”. Since the 1860s (Pancher and Deplanche) and even more recently, the habitat in the Babouillat area has been severely impacted by anthropogenic fires and invasive Rusa deer ( Rusa timorensis ), so this might explain its rarity or extinction, such as is the case for the Critically Endangered and possibly extinct Brunoniella neocaledonica ( Heine 1976: 34) Moylan (2000: 480) ( Tanguy et al. 2020). Since we (GG and GL), Dominique Fleurot and David Bruy have spent several days searching unsuccessfully for the species in the field, we can consider it as Extinct (EX), following IUCN (2019) guidelines. If the local Red List unit ( Meyer et al. 2022) confirms this assessment, it will be the second New Caledonian endemic species after Pycnandra micrantha ( Beauvisage 1901: 88) Munzinger & Swenson (2015: 102) to fall into this category.
Notes:—The material of Pytinicarpa tonitrui contains some specimens ( Deplanche 425, Mus . Néocal. 94 [ Pancher 94 / Vieillard 2823]) originally cited by Guillaumin (1937) as syntypes of Brachyscome neocaledonica , a name here put in the synonymy of P. sarasinii . Unfortunately, Wang et al. (2022) based the lectotypification of B. neocaledonica only on online scans, and failed to observe that two actually different species were mixed within syntypes of B. neocaledonica . Our careful examination of the original material revealed several distinctive characters ( Table 1 View TABLE 1 ). For instance, the main distinctive character is the pubescence of the ray florets corolla tube which is glabrous in P. tonitrui , and systematically hairy in P. sarasinii . Florets are also much shorter in P. tonitrui than in P. sarasinii , respectively 0.5–1 mm vs. 1–1.3 mm long for the ray florets, and ca 1.3 mm vs. ca. 2.5 mm long for the disc florets. As far as observed, P. sarasinii remains the only Pytinicarpa in New Caledonia to have hermaphrodite disc floret, all disc florets observed for P. tonitrui being functionally male. Further, sterile material can be differentiated between both species, as P. sarasinii are always hairy plants, while P. tonitrui tends to be almost glabrous, with the pubescence limited to the leaf base. Yet the distinction on sterile material remains tricky and needs to be confirmed on fertile material, the pubescence of the ray floret corolla tube being the final distinctive character. Consequently, within some sheets of Deplanche 425, a gathering originally cited as syntype of B. neocaledonica , some individual plants belong to P. tonitrui . P00537800 and K000890083 plants all belong to P. tonitrui . P00537799 is a mixed specimen, with the two plants on the left belonging to P. tonitrui , while the two on the right belong to P. sarasinii . P00537801 (said to be from Neue, not an original label) is clearly a mixed specimen with plants probably collected in different places and at different times. Some plants belong to P. sarasinii , which were most likely collected in the Koniambo area (Gatope s.l.), but it was impossible to definitively identify each plant.
From all the specimens known, only one Mus . Néocal.94 (P00537795), was retrieved and belongs to P. tonitrui , as confirmed by Guillaumin’s drawings showing the glabrous ray floret corolla tube.
Specimens examined:— NEW CALEDONIA. North Prov.Nené [Neué], 1861–1867, Deplanche 425 (K000890083image!, P00537799p.p.! see notes, P00537800!); Néué, 1870, Mus . Néocal. [J.A.I. Pancher] 94 [E. Vieillard 2823] (P00537795).

| H |
University of Helsinki |
| S |
Department of Botany, Swedish Museum of Natural History |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |