
Rhyacodrilus saelonae, Rodriguez, Pilar & Fend, Steven V., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3664.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C8136C89-7787-477D-BC64-97AA6C16057B |
|
DOI |
https://doi.org/10.5281/zenodo.6160649 |
|
persistent identifier |
https://treatment.plazi.org/id/154FD01D-4B51-FFD4-FF6E-92AFBBE3CBA8 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhyacodrilus saelonae |
| status |
sp. nov. |
Rhyacodrilus saelonae sp. n.
( Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Holotype. USNM 1202047, whole-mounted worm stained in hematoxylin and mounted in Canada balsam.
Paratypes. USNM 1202048-52: 3 mature specimens from the type locality, stained in hematoxylin or borax carmine and whole-mounted (17 March 2008); 1 mature, dissected worm, stained with hematoxylin (7 May 2006); 1 sagittally sectioned (17 March 2008). MNCN 16.03/3079: 3 mature specimens from the type locality, stained in borax carmine and whole-mounted (17 March 2008).
Type locality. Nacimiento River, above Fort Hunter Liggett, at upstream bridge, Nacimiento-Fergusson Road, Monterey Co., California, USA. N36.0132°, W121.4201° (17 March 2008).
Other material. From the type locality: 1 whole mount (22 April 2006), 8 whole mounts and 1 dissected (7 May 2006), 2 sagittally sectioned and 13 whole mounts (17 March 2008). Nacimiento River below Negro Fork, N35.9969° W121.3797°, 6 whole mounts (3 April 2008).
Etymology. The species is dedicated to Saelon Renkes, who has contributed to the discovery of many Nearctic oligochaete species.
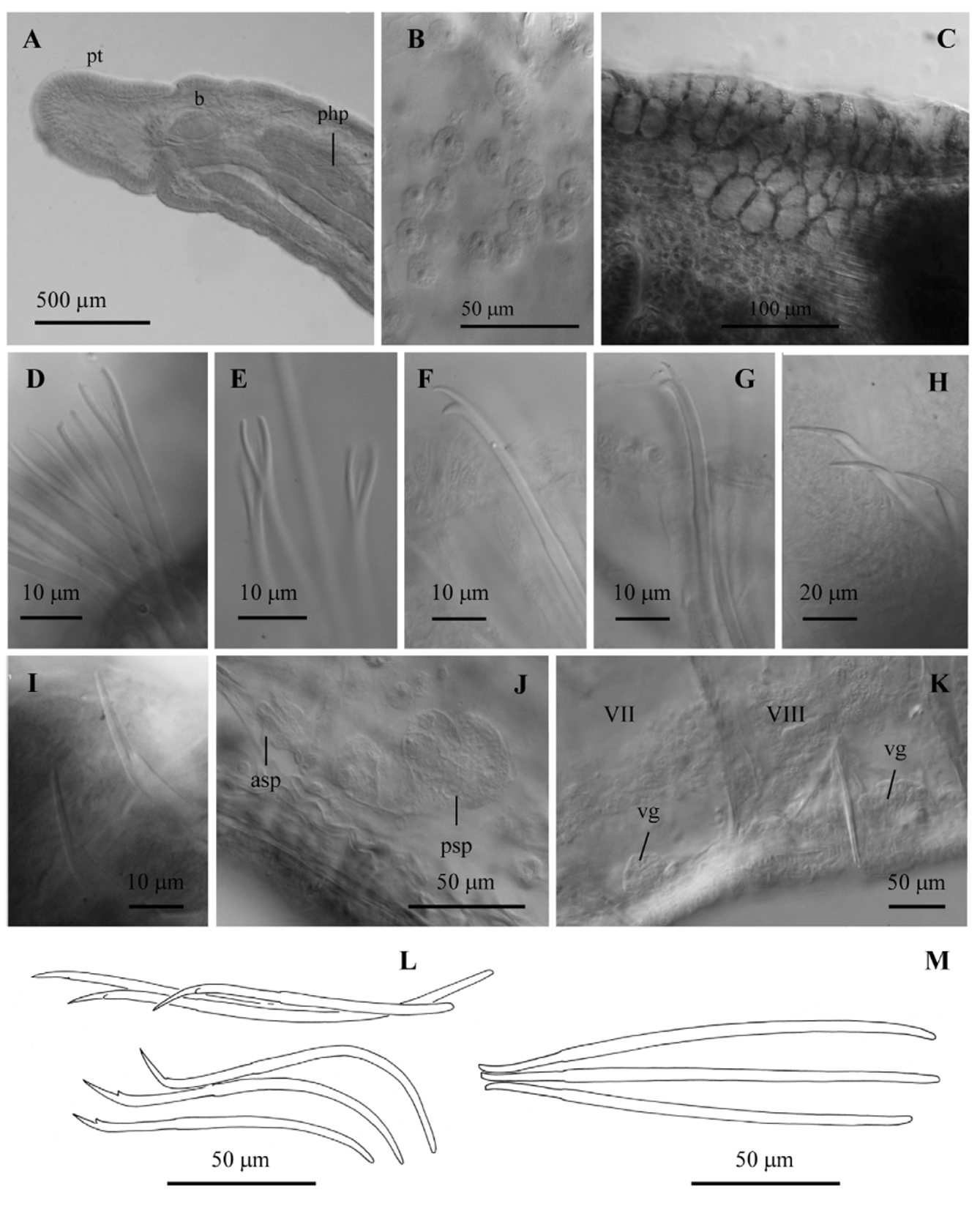
Description. (Measurements are based on individuals with sperm in spermathecae, with or without welldeveloped eggs). Number of segments 65–96 (based on 5 complete specimens). Diameter of the body in segment VIII (whole-mounted specimens) 217–362 µm. Elongated, round prostomium (197–240 µm long) ( Figs 1 View FIGURE 1 A, 2A). Secondary annulation clearly marked in II; faint but visible in other preclitellar segments of most specimens. Epidermis not densely glandular (8–12 µm high), small glandular cells unordered. Clitellum from the level of chaetae in the spermathecal segment to the end of the post-atrial segment, thickest dorsally and laterally, weak near male and spermathecal pores; clitellar epidermis up to 47 µm high (in most specimens about 20–30 μm), formed by large glandular cells in the dorsal and lateral part, extending ventrally in the post-atrial segment; ventral epidermis thickened but not glandular in the spermathecal segment, and thin in the area between the male pores ( Figs 1 View FIGURE 1 A, B, 2C).
Dorsal bundles composed of smooth hair-like and pectinate chaetae. Hair chaetae (1)2–3 per bundle anteriorly, and (0)1–2(3) in postclitellar segments, much longer than body diameter (408–800 µm) in the anterior segments, and longer or about the same as body diameter in posterior segments. Usually 3–4 pectinate chaetae in anterior bundles (rarely 2), and (1) 2–3 in postclitellar segments; 67–111 µm long (much shorter in II, down to 41µm). Pectinates in anterior segments with lyre-shaped lateral teeth (7–13 µm long) and 2–4 shorter (4–8 µm) intermediate teeth; pectinates in posterior segments with shorter lateral teeth (6–8 µm long) ( Fig. 2 View FIGURE 2 D, E). The last segments of complete specimens with only 1 pectinate chaeta (48 µm long) per bundle, with very short teeth, and without hair chaetae. Ventral chaetae bifid; 5–9 per bundle (rarely 3 in segment II) in anterior segments, 4–7 in postclitellar segments; distal tooth 1.6–1.8 times longer than proximal in anterior segments, becoming shorter (about same length) and thinner posteriorly ( Fig. 2 View FIGURE 2 F, G). In the last segments down to 1–3 ventral chaetae per bundle, with distal tooth shorter and thinner than proximal.
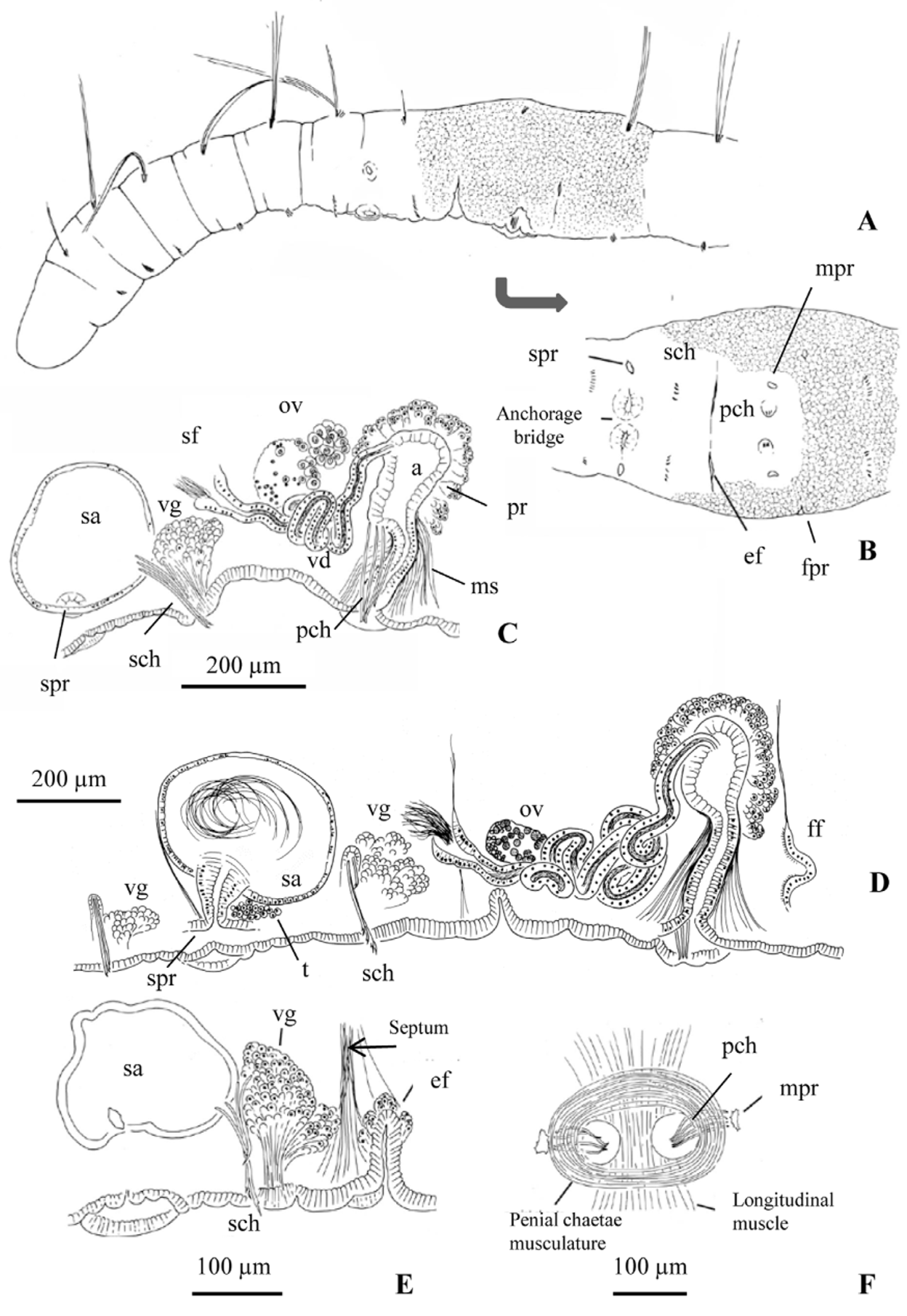
One pair spermathecal pores; simple, oval openings, up to midway between ventral chaetal line and lateral line, near anterior septum of the testicular segment. Modified spermathecal chaetae behind spermathecal pores, (3)4–5 per bundle (commonly, one of them developing), in line with ventral chaetae, sometimes on a low mound with cellular extensions from the ventral gland penetrating the epidermis behind chaetae. Spermathecal chaetae 116–158 µm long and 4 µm shaft diameter, with very long, sharply-pointed, grooved distal tooth (14–19 µm long); the short, proximal tooth appears as a wide prominence at the base ( Figs 2 View FIGURE 2 H, I, L). Nodulus 0.3–0.4 from the distal end. Two ridges of raised epidermis, U-shaped to nearly circular, open laterally (diameter 80–100 μm, 20–35 μm high at midline) and conjoined at ventral midline between the spermathecal pores, forming an "anchorage bridge" (sensu Cuadrado and Martínez-Ansemil, 2001). A conspicuous, transverse epidermal fold (about 80–120 μm wide, up to 30–50 μm deep) opens at septum behind (and in line with) spermathecal chaetae.
One pair simple male pores in the ovarian segment, on (or slightly median to) the ventral chaetal line, at about the posterior 1/4 of the segment. Modified penial chaetae slightly median to the male pore, in the atrial segment, single-pointed, 120–161 µm long, 3–4 per bundle, weak nodulus at 0.2 from distal end ( Fig. 2 View FIGURE 2 M). Penial chaetae all of the same size or the middle ones slightly longer; arranged roughly fanwise, with tips close together and directed medially; usually on low tubercles (about 25 µm high); the tubercles and chaetae surrounded by a ring of muscles ( Fig. 1 View FIGURE 1 F). A complex of dorso-ventral muscular strands is also associated with the penial chaetae, attaching to the tubercle near the tips of the chaetae; bases of chaetae have the usual chaetal musculature attaching them to the ventral body wall. One pair female pores at posterior intersegment of ovarian segment, midway between ventrolateral chaetal line and lateral line. Reproductive organs are often shifted forward by 1 or 2 segments relative to typical Rhyacodrilus , but they retain their relative positions: spermathecal pores in VIII, IX, or X correspond to male pores in IX, X, or XI, respectively, and the clitellum begins in the spermathecal segment.
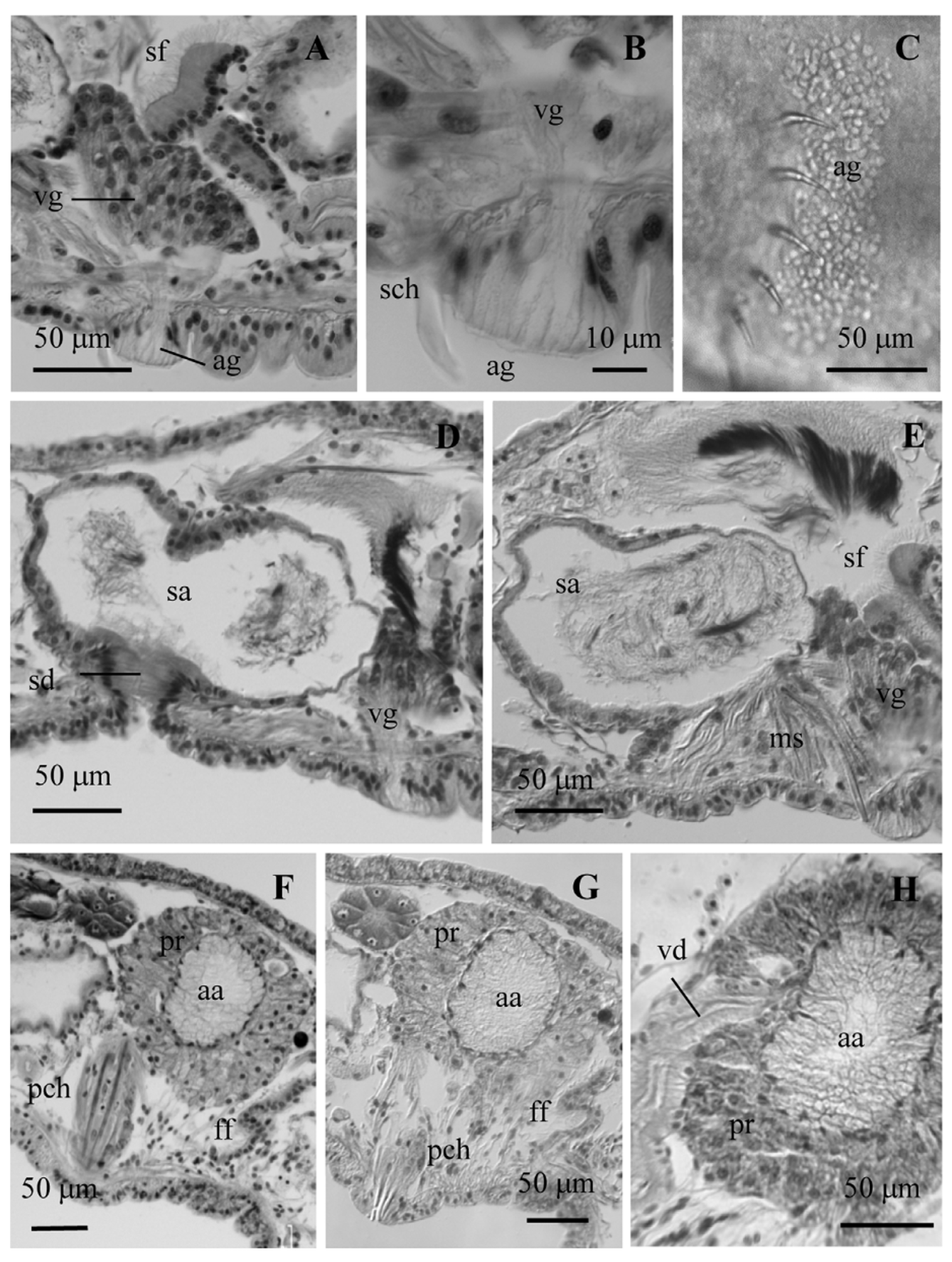
Nucleated coelomocytes with granulated cytoplasm (diameter 12–18 µm) ( Fig. 2 View FIGURE 2 B). Pharyngeal glands welldeveloped in IV–V, usually extending into VI. Nephridia paired, single, or absent in the postclitellar segments, sometimes in one or more preclitellar segments; with short, narrow anteseptal funnel, an elongate (sometimes inflated), granular postseptale ( Fig. 2 View FIGURE 2 J), and long efferent duct which may extend posteriorly into the next segment; the duct terminates at an inconspicuous nephropore just in front of the ventral chaetae. One pair ventral glands behind the ventral bundles of chaetae, from segment (IV)V to the spermathecal segment, and from the post-atrial segment as far back as XX; the largest in the spermathecal segment (up to 140 μm wide), well-developed in prespermathecal and post-atrial segments (50–90 μm wide), then progressively smaller. These glands project freely into the coelomic cavity and the cell necks extend through the muscular layers and body wall to the body surface forming an area glareosa (sensu Schmelz 2003: 49f) visible in vivo ( Figs 1 View FIGURE 1 E, 2K, 3A–C ag). Sperm sac may extend anteriorly to segments VIII and IX, and backwards to segment XIII. Egg sac may extend back to segment XIV.
One pair testes in the spermathecal segment (typically IX) and one pair ovaries in the atrial segment (typically X). Spermathecae composed of a short, conical duct, usually shorter than wide (34–60 µm long, 48–62 µm maximum diameter), with a thick wall of columnar epithelium, and an oval ampulla which occupies most of the length of the segment (120–282 µm maximum diameter in mated worms), with thin epithelium (6–12 µm high). Sperm free within lumen of spermathecal ampulla, but anterior ends of spermatozoids may be oriented towards one end of the ampulla. One ampulla may penetrate into an adjacent segment. Several specimens show in lateral view a dorsal fold in the ampulla ( Fig. 3 View FIGURE 3 D).
Male duct opens into a conical sperm funnel; vas deferens highly convoluted, ciliated, and of nearly uniform width (diameter 24–37 µm), length 530–750 μm, running along the ventral side of the segment and narrowing as it joins the atrium subapically, or in some specimens near the middle of the ampulla. Atrial ampulla ovate (95–175 µm long, 54–96 μm wide), completely covered by a thick (30–60 μm) layer of prostate cells, at least some of which are in densely packed clusters of 10 or more cells. Ampullar epithelium appears irregular and vacuolated, to 25 μm thick, sometimes nearly filling the atrial lumen ( Fig. 3 View FIGURE 3 F–H). Atrial duct 100–194 µm long, 43–52 µm wide), slightly narrowed entally, with a uniform, columnar epithelium, narrow lumen, and thin outer muscle layer; a dense array of muscular strands runs from ventral body wall to the outer muscle layer along the entire length of the duct ( Fig. 1 View FIGURE 1 C, D). Female funnels long, in some specimens more than half the diameter of the body ( Fig. 3 View FIGURE 3 F, G).
Anomalies. Positions of reproductive organs vary considerably in this species. More than half of the specimens had the genitalia shifted one or two segments forward from the usual positions in the Rhyacodrilinae . Of 36 mature worms examined, 12 had the spermathecae in X, 16 in IX, and 8 in VIII; in all cases the relative position of other reproductive organs to the spermathecae was preserved, and both male ducts and spermathecae were functional (with sperm). Some species of the genus Rhyacodrilus have been reported to have architomy followed by regeneration, which may be associated with a shift in the position of the reproductive segments (e.g. Snimschikova 1985), although we have not seen any evidence of developing anterior segments in this species. Two specimens had enlarged ventral chaetae in the post-atrial segment with upper teeth much longer than lower, similar to the spermathecal chaetae (but smaller); in this same segment there was also a small gonad and a slightly enlarged ventral gland behind the chaetal bundle. One specimen had an extra spermatheca on one side of the body, at the atrial segment (X), but only the spermatheca in the preatrial segment was filled with sperm. Another specimen had a single spermatheca (with sperm) in VII, a rudimentary spermatheca in VI, one pair spermathecae with a single functional male duct in VIII, and functional male ducts paired in IX.
Distribution and habitat. Known only from type locality, a largely undisturbed section of the Nacimiento River, in the Santa Lucia Mountains (Los Padres National Forest). The two reaches with R. saelonae were in a small stream with permanent surface flow, near the transition from high to low gradient sections. Sediment was a mixture of colluvial and alluvial particles, sizes ranging from cobbles to coarse gravels, overlaying coarse sand. The sampling area is a salmonid habitat, with clear, cool water, a well-preserved riparian habitat and little aquatic vegetation.
Remarks. Based on the relative position of the reproductive organs (spermatheca in the most anterior part of the testicular segment, plesioporous male duct and male pores opening in the ovarian segment), the prostate diffuse or with cells in clusters broadly attached to the atrial surface (Erséus 1984), and the abundance of large nucleated and granulated coelomocytes, the new species is member of the subfamily Rhyacodrilinae . Although this subfamily requires revision due to its paraphyletic nature (Erséus et al. 2008, Martin et al. 2010), the new species has all the main diagnostic characteristics of the genus Rhyacodrilus : moderately long vasa deferentia entering atria subapically, well-developed prostate glands covering the atrial ampulla, coelomocytes present, and modified penial chaetae (Brinkhurst & Jamieson 1971).
The group of Rhyacodrilus species having both spermathecal and penial chaetae is summarized in Table 1. Among these species, only two were previously known that share both hair and pectinate chaetae in dorsal bundles and a well-developed layer of prostate cells: R. svetlovi Sokolskaya, 1976 (Chukchi Peninsula, Siberia) and R. tauricus Dembitsky, 1975 (Crimea, Ukraine). The former differs from R. saelonae sp. n. in several respects: It lacks hair chaetae in segments posterior to XV, the atria are tubular, the vas deferens is shorter than the atrium, the atrial duct is very short, and the vas deferens joins the atrium apically. The new species is separated from R. tauricus by the shape of the spermathecal and posterior dorsal chaetae as well as by the junction of vas deferens, which is on the posterior face of the atrium in R. tauricus . Modified spermathecal-like chaetae are present in both preatrial and postatrial segments in R. tauricus . The general structure of the male duct in R. saelonae sp. n. (i.e. atrial duct as long as or longer than atrial ampulla, and long vas deferens entering atrium subapically), together with the presence of both penial and spermathecal chaetae, the extraordinarily long distal tooth of the spermathecal chaetae, the lateral position of spermathecal pores, and the presence of ventral glands clearly distinguish this species within the genus Rhyacodrilus .
The ventral glands in R. saelonae sp. n. are similar in structure to Timm's glands, described in Haplotaxidae by Hrabĕ (1931), and are another diagnostic character for the new species. In our collection, partially mature R. saelonae have small glands in the spermathecal segment which are similar in size to the ventral glands in other anterior segments (based on the examination of 8 specimens at varying stages of reproductive development). The glands in the spermathecal segment are well-developed (larger than those in adjacent segments) in two mature but unmated specimens. This development of the glands in the spermathecal segment during maturation, as well as their position close to the spermathecal chaetae, suggests some role during mating.
The presence of modified chaetae in the postatrial segment is extremely rare among rhyacodrilines, although they have been reported in Protuberodrilus Giani and Martínez-Ansemil, 1979 and Rhyacodrilus tauricus . In R. saelonae sp. n., this occurred in only two specimens and is interpreted as an anomaly.
Among Rhyacodrilus species, a lateral position of the spermathecal pores has been reported for R. gasparoi Martínez-Ansemil et al., 1997 , a cave species from Italy. That species also has well-developed ventral glands in segment X, associated with spermathecal chaetae. The distinct transverse folds between the spermathecal and atrial segments of the new species correspond to the position of the spermathecal chaetae in the expected inverted copulatory orientation, and thus may have a copulatory function. Other external copulatory structures of R. saelonae appear to be a variation on those described by Cuadrado and Martínez-Ansemil (2001) for Peristodrilus montanus (Hrabĕ, 1962) . The relative positions of the anchorage bridge (here formed by two concave tubercles), the penial chaetae, and male and spermathecal pores are similar. The low tubercles and dorso-ventral muscles associated with penial chaetae may correspond to the "raised ventral elevation" and associated musculature described by Cuadrado and Martínez-Ansemil (2001) for P. montanus . The ring of circular muscles surrounding the penial chaetae in R. saelonae was not mentioned in the latter account, but would have the function of pulling the tips of the chaetae (and the tubercles) together. As the bases of these chaetae are fixed by normal chaetal musculature, the effect would be forceps-like.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |