
Balsateres, González-Santillán & Prendini, 2013
|
publication ID |
https://doi.org/10.1206/830.1 |
|
DOI |
https://doi.org/10.5281/zenodo.4624293 |
|
persistent identifier |
https://treatment.plazi.org/id/174CE445-FFE6-2E48-0B39-9C0196A7FA65 |
|
treatment provided by |
Felipe |
|
scientific name |
Balsateres |
| status |
gen. nov. |
Balsateres View in CoL , gen. nov.
Figures 3 View Fig , 7 View Fig , 16A View Fig , 20A View Fig , 28A View Fig ; table 1 View TABLE 1
Vaejovis cisnerosi Ponce-Saavedra and Sissom, 2004 [ = Balsateres cisnerosi ( Ponce-Saavedra and Sissom, 2004) View in CoL , comb. nov.], type species, by monotypy.
Thorellius View in CoL (part): Soleglad and Fet, 2008: 1 View Cited Treatment , 95, 102; Ayrey and Soleglad, 2011: 1.
Vaejovis intrepidus group (part): Santibáñez-López and Sissom, 2010: 52.
ETYMOLOGY: The name Balsateres , gen. nov., a compound word, masculine in gender, is derived from the Balsas Depression, to which the genus is endemic, and the Latin adjective teres (‘‘smooth’’ or ‘‘polished’’) which refers to the glabrous integument typical of this genus.
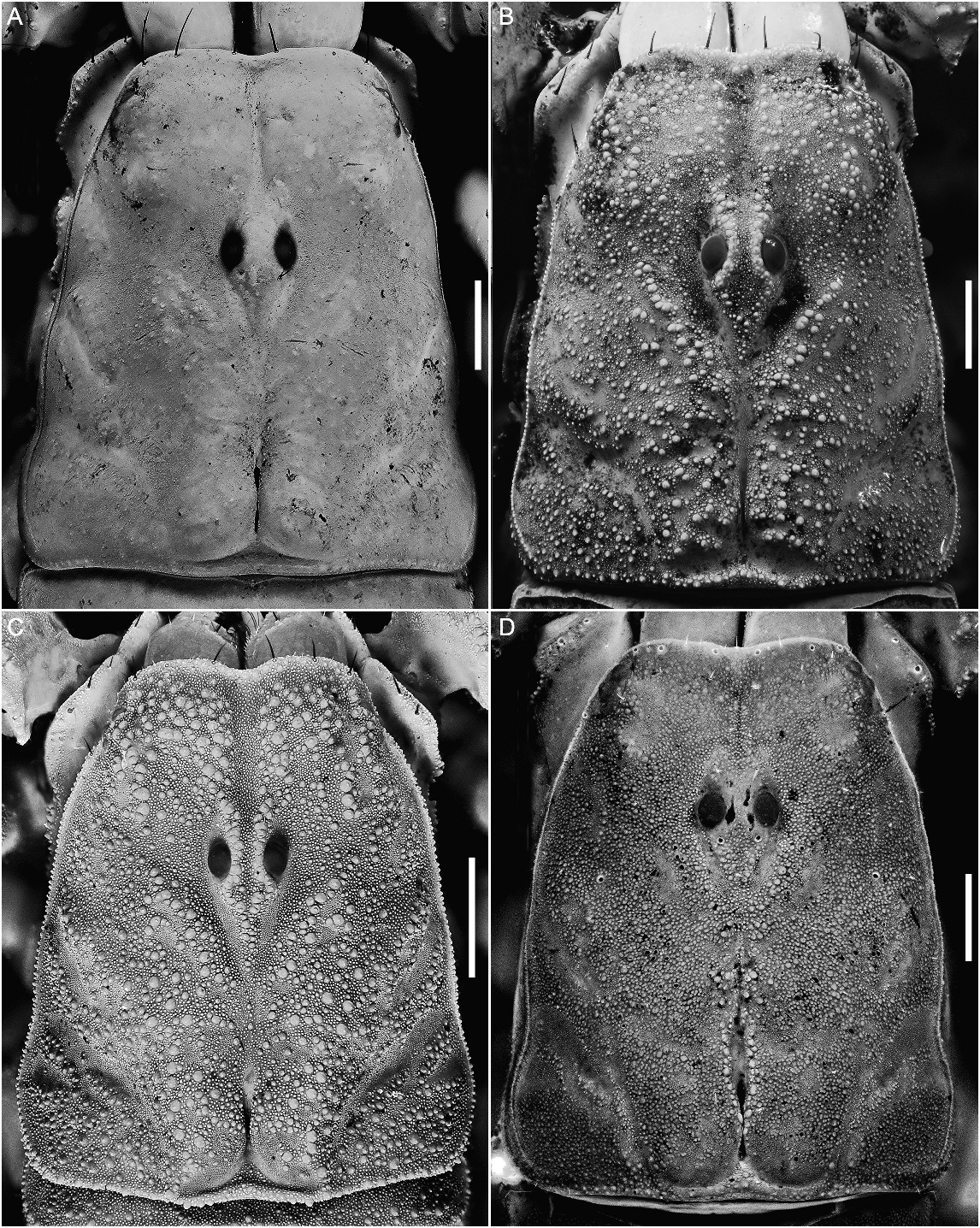
DIAGNOSIS: Balsateres , gen. nov., is distinguished from other genera of Syntropinae by the smooth dl carinae on metasomal segments I–IV. The carinae and intercarinal surfaces of the carapace (fig. 16A), pedipalp chela, patella, tergites, metasoma, and telson (fig. 28A) are smooth in this genus. Counts of macrosetae on the carinae of metasomal segments I–IV are also greatly reduced, as follows: dl, 0/0:0/0:0/0:1/1:4/4; lm, 0/0:0/0:0/ 0:0/0:2/2; vl and vsm, 1/1:1/1:1/1:1/1:3/3. Two or more macrosetae are present on these carinae in all other Syntropinae , including Thorellius , which also presents low setal counts, e.g., Thorellius cristimanus (Pocock, 1989) with the following counts on segments I– V: dl, 2/2:3/3:3/3:3/3:7/6; lm, 1/1:3/3:3/3:4/4:4/4; vl, 2/2:3/3:3/3:3/3:7/7; vsm, 3/3:3/3:3/3:3/3:5/5.
Balsateres , gen. nov., is most closely related to Thorellius (fig. 7), with which it shares broad pedipalpal and metasomal carinae and, with Thorellius intrepidus (Thorell, 1867) , double basal prolateral denticles on the movable finger of the pedipalp chela (fig. 20A, B). However, all carinae are smooth in Balsateres , gen. nov., instead of moderately to densely granular, as in Thorellius . The distinct proximal gap, evident when the pedipalp chela fingers are closed, in both sexes of Thorellius , is absent in Balsateres , gen. nov. The two genera also differ in base coloration and infuscation. Balsateres , gen. nov., is yellowish and almost immaculate, except for the ocular tubercle, which is outlined by dark infuscation, whereas Thorellius tend to be darker and reddish in color, with various degrees of infuscation on the carapace and tergites.
Balsateres , gen. nov., resembles Mesomexovis , gen. nov., in possessing obsolete ventral carinae on metasomal segments I– IV, but species of the latter genus differ in being more setose and infuscate.
INCLUDED SPECIES: Balsateres cisnerosi ( Ponce-Saavedra and Sissom, 2004) , comb. nov.
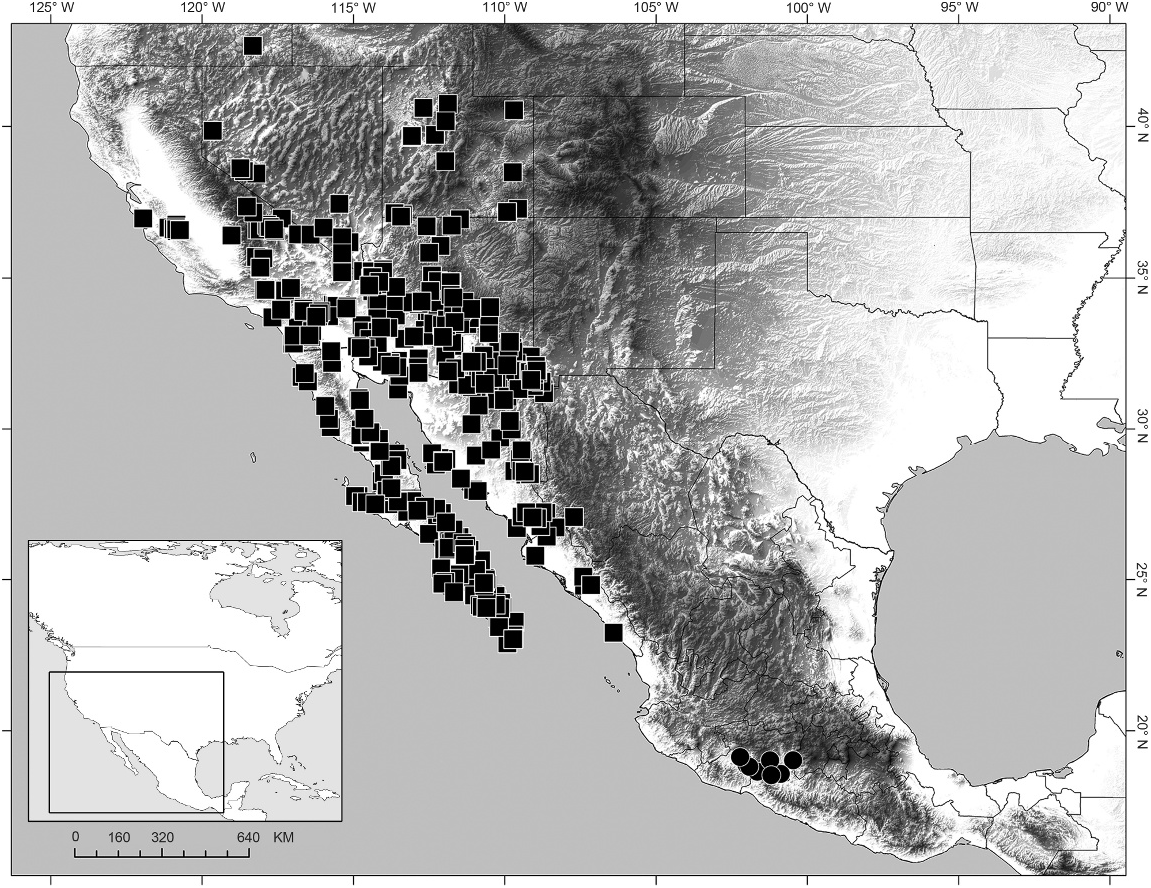
DISTRIBUTION: Balsateres , gen. nov., is endemic to the Balsas Depression of Mexico and recorded only from Estado de México and Michoacán (fig. 3).
NATURAL HISTORY: Balsateres , gen. nov., inhabits tropical deciduous forest at altitudes between 195– 672 m. Specimens were observed at night, doorkeeping at the entrances of their burrows in open, sandy areas, and excavated from the burrows ( Ponce-Saavedra and Sissom, 2004). The glabrous integument and burrowing biology in sandy habitats suggest that this species is psammophilous although the leg setation is atypical for the psammophilous ecomorphotype, because the dorsal and retrolateral dorsal macrosetae are arranged into two separate rows, not forming a setal comb. This monotypic genus was collected at the same locality as another monotypic syntropine genus, Kuarapu . Although sympatric, these genera are allotopic: Balsateres , gen. nov., was collected in open, sandy areas, whereas Kuarapu was collected on road-cuts, suggesting a lithophilous ecomorphotype.
REMARKS: Due to the unique morphology of this species, Ponce-Saavedra and Sissom (2004) refrained from assigning it to one of the five species groups of Vaejovis recognized at the time. Soleglad and Fet (2008) transferred it to Thorellius , a name devised for Hoffmann’s (1931) ‘‘second section’’ of Vaejovis , later termed the intrepidus group by Sissom (1989), without quantitatively testing its monophyly or composition. Santibáñez-López and Sissom (2010) also referred this species to the intrepidus group. The creation of a monotypic genus is merited by the unique diagnostic character combination of this species.
MATERIAL EXAMINED: Balsateres cisnerosi ( Ponce-Saavedra and Sissom, 2004) , comb. nov.: MEXICO: Michoacán: Municipio de La Huacana: El Vado , 18 ° 48.908 ′ N 101 ° 54.976 ′ W, 198 m, 20.v.2007, O.F. Francke, J. Ponce, M. Villaseñor, and A. Quijano, 1♂, 1♀ ( IBUNAM).



| IBUNAM |
Instituto de BiIología, Universidad Nacional Autónoma de México |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Syntropinae |
Balsateres
| González-Santillán, Edmundo & Prendini, Lorenzo 2013 |
Vaejovis intrepidus
| Santibanez-Lopez, C. & W. D. Sissom 2010: 52 |
Vaejovis cisnerosi
| Ponce-Saavedra, J. & W. D. Sissom 2004: 21 |