
Lygodactylus nyaneka, Marques & Ceríaco & Buehler & Bandei- Ra & Janota & Bauer, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4853.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:802D1C38-1186-4187-8289-5AC125242802 |
|
DOI |
https://doi.org/10.5281/zenodo.4506723 |
|
persistent identifier |
https://treatment.plazi.org/id/6CCB79F1-AE14-4CDB-8CAC-365C342ABBC5 |
|
taxon LSID |
lsid:zoobank.org:act:6CCB79F1-AE14-4CDB-8CAC-365C342ABBC5 |
|
treatment provided by |
Plazi |
|
scientific name |
Lygodactylus nyaneka |
| status |
sp. nov. |
Lygodactylus nyaneka View in CoL sp. nov.
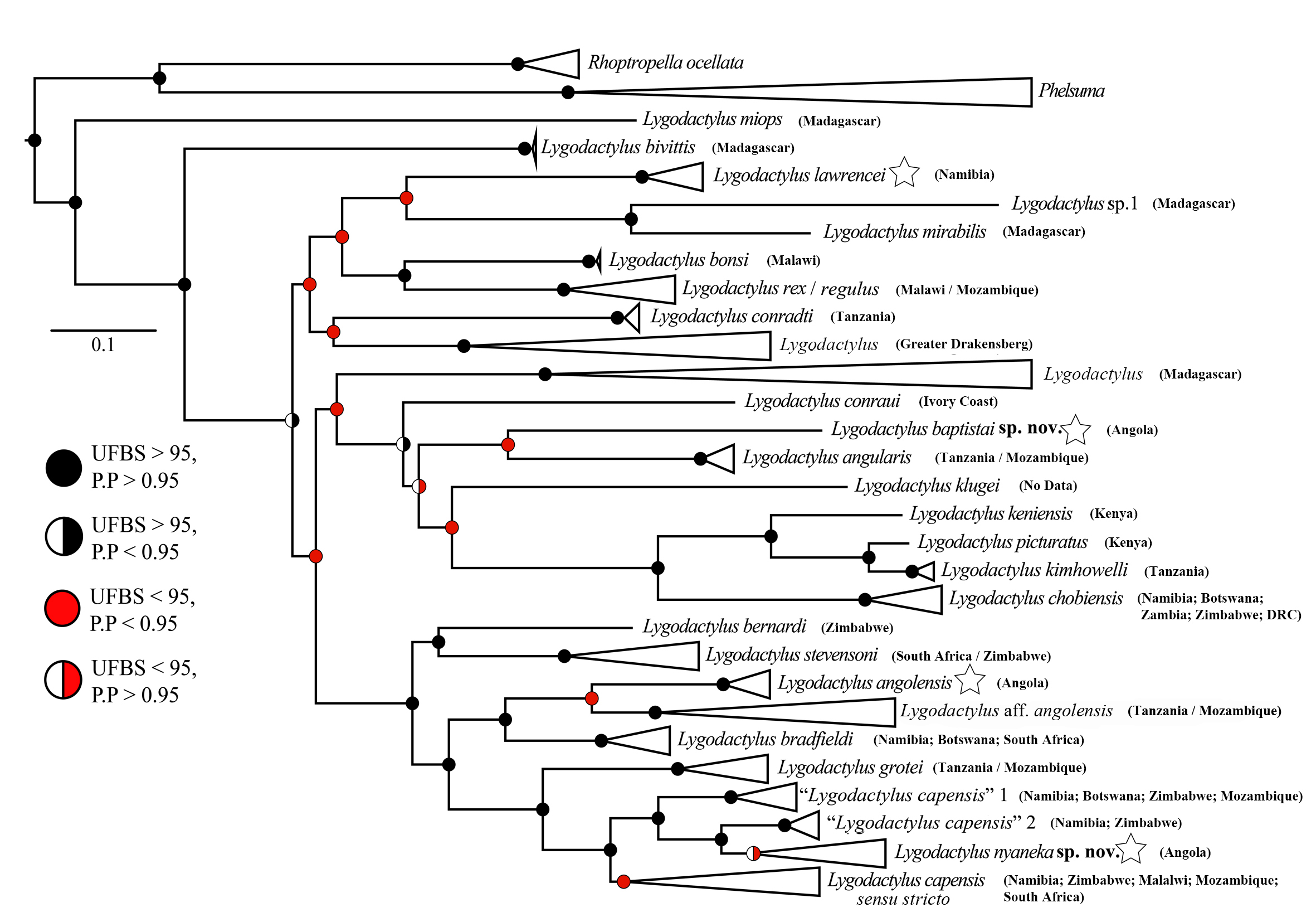
( Tables 4, 6; Fig. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 14–17 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 )
urn:lsid:zoobank.org:act:
Hemidactylus capensis View in CoL [sic] ( Bocage 1867: 219, 1870: 68)
Lygodactylus capensis View in CoL ( Bocage 1895: 15; Monard 1937: 53; Loveridge 1947: 208, 1957: 187; Frade 1963: 253; Pasteur 1965 [1964]: 62; Branch 1998: 246; Marques et al. 2018: 190)
Lygodactylus capensis capensis ( Schmidt 1933: 4; Bates et al. 2014: 118)
Lygodactylus capensis bradfieldi ( FitzSimons 1943: 53)
Lygodactylus bradfieldi View in CoL [part] ( Branch 1998: 246; Conradie et al. 2016: 24 View Cited Treatment [?]; Marques et al. 2018: 189,190)
Lygodactylus angolensis ( Baptista et al. 2019: 113) View in CoL View Cited Treatment
The majority of the authors who historically dealt with Angolan Lygodactylus other than L. angolensis considered them to represent L. capensis ( Bocage 1867, 1870, 1895; Schmidt 1933; Monard 1937; FitzSimons 1943; Loveridge 1947; Frade 1963; Pasteur 1965 [1964]; Branch 1998; Marques et al. 2018). Bocage (1867, 1895, 1896) recorded them from Namibe and Huíla provinces; Schmidt (1933) recorded them from Benguela Province; Monard (1937) recorded the species from Huila and Cunene provinces; Loveridge (1947) recorded them from Namibe Province; and Laurent (1964) recorded the species from Lunda Sul and Moxico provinces. Based on these records, the species was considered to occur in Angola by all authors who reviewed the genus in Angola ( Loveridge 1947; Pasteur 1965 [1964]; Branch 1998; Marques et al. 2018; Branch et al. 2019). However, Branch (1998) stated the possibility that besides L. capensis , the morphologically similar L. bradfieldi could also occur in Angola. Conradie et al. (2016) provided the first records of L. bradfieldi in the country based on specimens collected in Cuando Cubango province, and Marques et al. (2018) interpreted many of the historical records assigned to L. capensis as L. bradfieldi . Originally described by Hewitt (1932 a) based on specimens from near Okahandja in central Namibia, L. bradfieldi has been recorded from numerous places in Namibia, Angola, Botswana, Zambia Zimbabwe and South Africa (Mertens 1971; FitzSimons 1943; Loveridge 1947; Auerbach 1987; Broadley 1991; Jacobsen 2011; Bates et al. 2014; Marques et al. 2018). Morphologically the species is very similar to L. capensis , and most of the characters that are used to distinguish the species overlap considerably, making it difficult to easily diagnose the species from one another. This morphological similarity led some authors to consider the species as western race or a synonym of L. capensis ( FitzSimons 1943; Loveridge 1947, 1957), until Pasteur (1965 [1964]) resurrected it. According to Pasteur (1965 [1964]), the most useful characters to differentiate L. bradfieldi from L. capensis are the type of contact between the rostral and the naso-rostrals (rostral dividing the naso-rostrals in L. bradfieldi versus rostral not dividing the naso-rostrals in L. capensis ); a lower number of mid-dorsal scales ( 156–177 in L. bradfieldi versus 170–274 in L. capensis ); and staggered rows of subcaudal plates in original tails of L. bradfieldi , versus a more linear distribution of the rows in L. capensis . FitzSimons (1943) considered that the main distinguishing character between the taxa was the number of scales entering the nostril, which in L. bradfieldi is four (rostral, 1 st supralabial and two nasal scales) and in L. capensis is five (rostral, 1 st supralabial and three nasal scales). Following Pasteur (1965 [1964]), Broadley (1991), Branch (1998) and Jacobsen (2011) considered the arrangement of the subcaudal scales of original tails as a good character to differentiate between the species. Our results show that, contrary to earlier interpretations, the central and southwestern Angolan populations of Lygodactylus belong neither to typical L. capensis nor to true L. bradfieldi but to another of several clades within the L. capensis species complex. The type locality of L. capensis is not very precise (“Kaffirland, and the districts to the north of Cape Colony” Smith 1849), and the type material is lost fide FitzSimons (1937). Kaffirland, or Kaffraria, referred to southeastern regions of the Eastern Cape, where the species is presumably not native, but has been introduced ( Rebelo et al. 2019), while the “distrcits to the north of Cape Colony” was used by Smith for areas currently part of the Northern Cape Province and/or areas eastward to modern Gauteng. It is, therefore, safe to assume that the nominotypical form belongs to the South African clade ( Lygodactylus capensis 3 in Figure 1 View FIGURE 1 ; Table 2) and cannot be applied to the Angolan clade. Besides the original nomen, there are a few names in the synonymy of L. capensis but none apply to the Angolan populations, and therefore, the Angolan clade of L. capensis species complex remains an unnamed and undescribed taxon. A further detailed taxonomic revision of the L. capensis species complex is underway (S. Travers et al., unpublished). The preliminary identification of L. bradfieldi in Cuando Cubango by Conradie et al. (2016) and the interpretation by Marques et al. (2018) who considered the records of L. capensis from Namibe Province to belong to L. bradfieldi , based on the latter species’ distribution and adaptation to arid savanna habitats, is understandable given the subtle morphological differentiation between L. capensis species complex and L. bradfieldi . A review of our newly collected specimens and those historical museum specimens recorded as L. capensis for Angola revealed that all of these specimens have the subcaudal scales arrangement typical of L. bradfieldi and/or three nasal scales around the nostril (an L. capensis character). However, the molecular data provide evidence of the distinctiveness of these specimens from all other taxa, and morphological data also allow us to separate them from L. capensis sensu stricto and L. bradfiedli sensu stricto Besides the historical data, the new taxon has been found in Namibe and Huíla provinces (this paper; see Specimens examined below). Specimens cited by Bocage (1867, 1895) from Benguela and Huíla provinces were not analyzed, as they were lost in the fire that destroyed the Lisbon museum in 1978. However, the distribution of the species, as well as the limited data provided by Bocage (1895, 1896) allow us to confidentially allocate these specimens to the new taxon. The specimens from Cuando Cubango cited by Conradie et al. (2016) need molecular and morphological reevaluation.
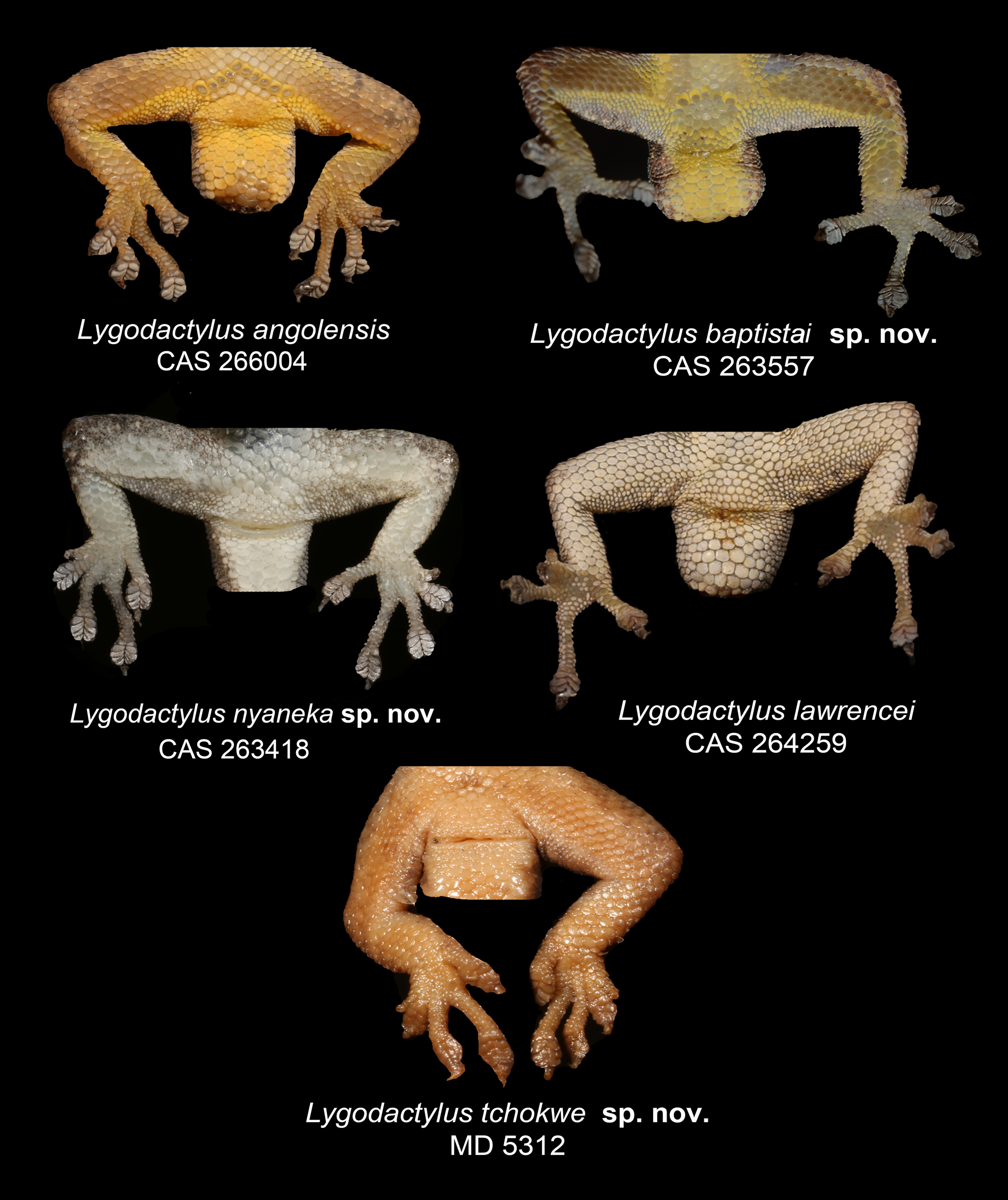
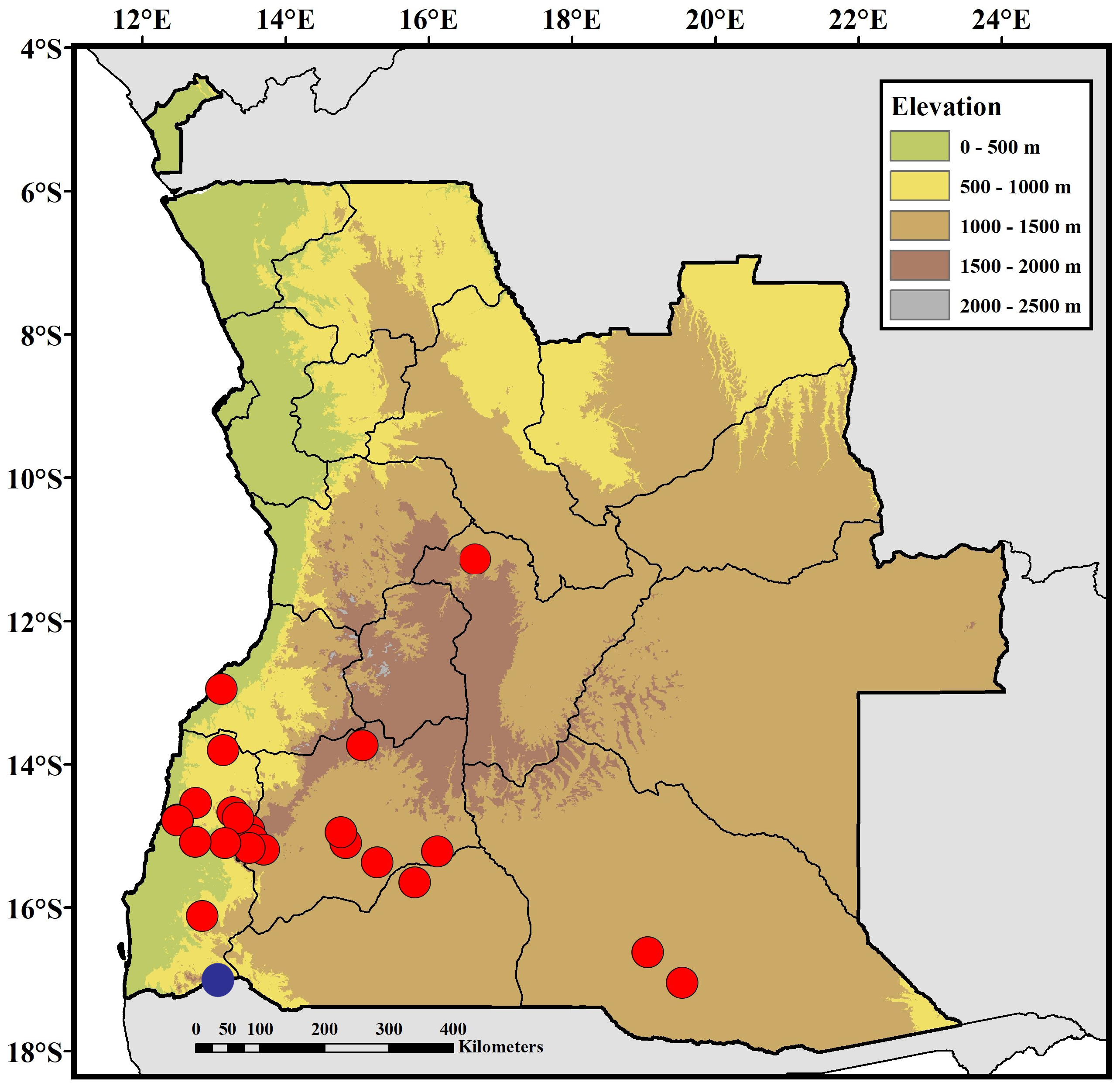
Holotype. An adult male ( CAS 263418 About CAS , field number AMB 10134; Fig. 14 View FIGURE 14 ) collected at Lubango, Kimbo do Soba, Huíla Province, southwestern Angola [-14.93419º, 13.46925º, 1866 m a.s.l], collected by M. Marques, L. Ceríaco, B. Butler, T. Júlio and M. Heinicke, on 21 July 2017.
Paratypes. All specimens from Angola. Six specimens: An adult female ( CAS 263420 About CAS , field number AMB 10136), same data as holotype ; An adult male ( CAS 263529 About CAS , field number AMB 10363), collected at Bibala Crossroad [-14.74350º, 13.33222º, 864 m a.s.l.], collected by L. Ceríaco, S. Bandeira & I. Agarwal, on 23 November 2016 ; An adult male ( CAS 263531 About CAS , field number AMB 10352), collected at Montipa [-14.66482º, 13.25763º, 669 m a.s.l.], collected by L. Ceríaco, S. Bandeira & I. Agarwal, on 23 November 2016 ; An adult male ( UF 187228, field number AMB 10165), collected at Bicuar National Park , camp headquarters [-15.10161º, 14.83986º, 1242 m a.s.l.], collected by M. Marques, L. Ceríaco, B. Butler, S. Bandeira, T. Júlio and M. Heinicke, on 27 July 2017 ; An adult male ( UF 187229, field number AMB 10790), collected at Chibia [-15.18944º, 13.69058º, 1467 m a.s.l.], collected by M. Marques, L. Ceríaco, B. Butler, S. Bandeira, T. Júlio and M. Heinicke, on 5 August 2017 ; An adult male ( UF 187230, field number AMB 10477), collected at Virei [-16.11958º, 12.83458524º, 523 m a.s.l.], collected by L. Ceríaco, S. Bandeira & I. Agarwal, on 28 November 2016 .
Additional material. ANGOLA: Bié Province: 14 km by dirt road from Cassumbi village , in a banana plantation [-11.08391º, 16.66367º, 1240 m a.s.l.] ( MHNC-UP / REP 499 ) . Huíla Province: Lubango, Kimbo do Soba [-14.93419º, 13.46925º, 1866 m a.s.l] ( CAS 263417 About CAS , 263419 About CAS , 263421 About CAS ; UF 187226; INBAC / AMB 10133, 10137 View Materials , 10134 View Materials ) ; Lubango, Instituto Superior de Ciências da Educação ( ISCED) [-14.91822º, 13.48492º, 1795 m a.s.l.] ( CAS 263422 About CAS ) ; Huíla waterfall [-15.05488º, 13.53491º, 1745 m a.s.l.] ( CAS 263405 About CAS ) ; Bicuar National Park , camp headquarters [-15.10161º, 14.83986º, 1242 m a.s.l.] ( UF 187227) ; Bicuar National Park, Hombo entrance [-14.94714º, 14.76947º, 1307 m a.s.l.] ( MHNCUP /Rep 435) ; Bicuar National Park, Matunto entrance [-15.36935º, 15.27510º, 1157 m a.s.l.] ( MHNCUP / REP 436–7 ) ; Kampulu (= Campulu-Cambissa) [-15.21667º, 16.11667º, 1268 m a.s.l.] ( MHNC 91.0445 View Materials ) . Namibe Province: Montipa [-14.66482º, 13.25763º, 669 m a.s.l.] ( CAS 263530 About CAS , 263532–33 About CAS ) ; Bibala Crossroad [-14.74350º, 13.33222º, 864 m a.s.l.] ( CAS 263525–28 About CAS ; UF 187232; INBAC / AMB 10362) ; Virei [-16.11958º, 12.83458524º, 523 m a.s.l.] ( CAS 263511–14 About CAS ; UF 187231) ; Mamué [-13.80080º, 13.12350º, 715 m a.s.l.] ( CAS 263556 About CAS ) ; Bumbo farm [-15.1678692º, 13.1491319º, 738 m a.s.l.] ( CAS 264242 About CAS ) ; Mucungu Farm [-14.77083º, 12.49231º, 333 m a.s.l.] ( CAS 264231 About CAS , 264232 About CAS ) ; between Ouipaca and Otchifengo [-16.63526º, 12.9246º, 598 m a.s.l.] ( MHNC-UP / REP 571 ) . Undetermined: Angola ( MHNC 91.0447 View Materials – 48 View Materials ) .
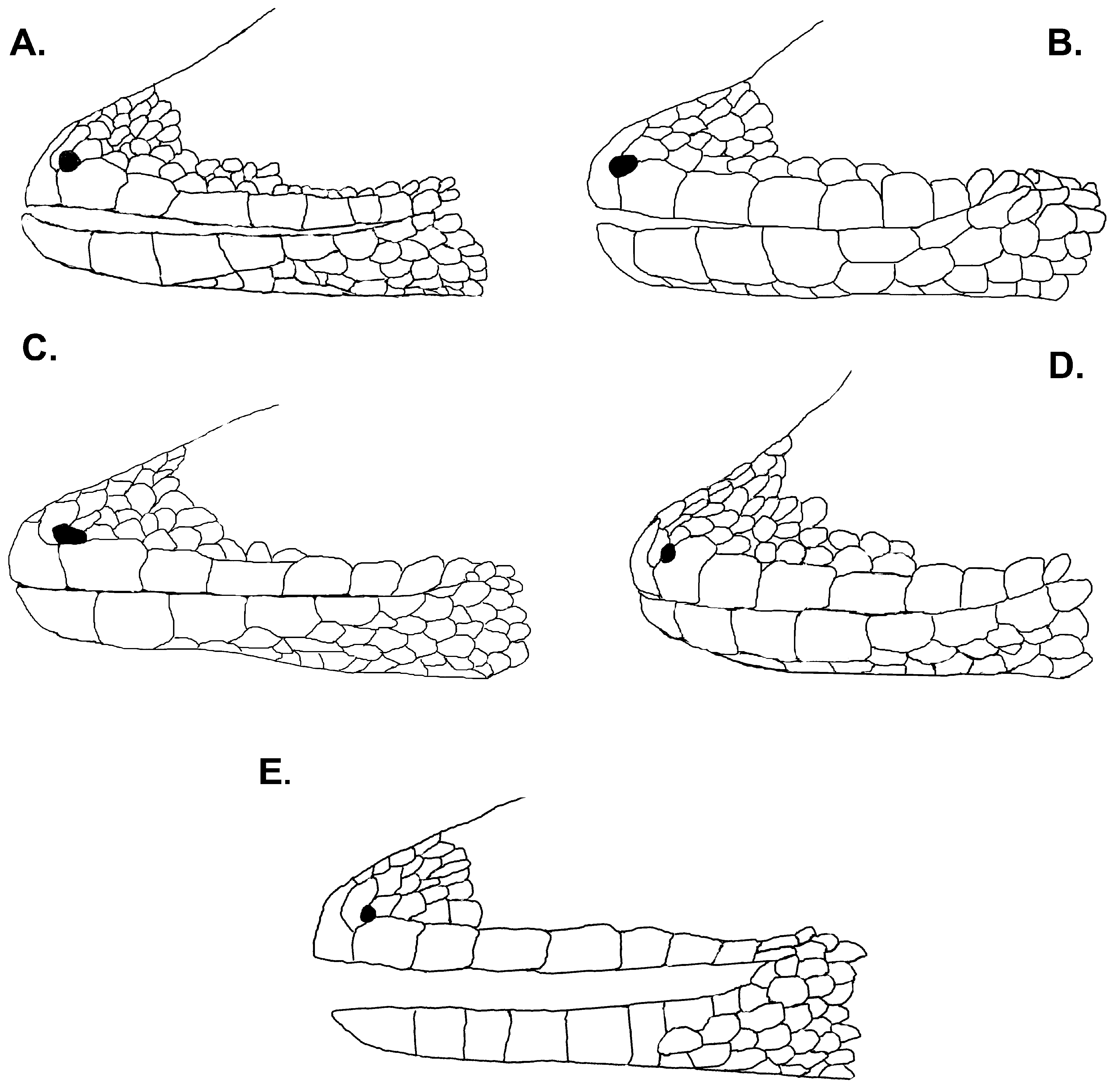
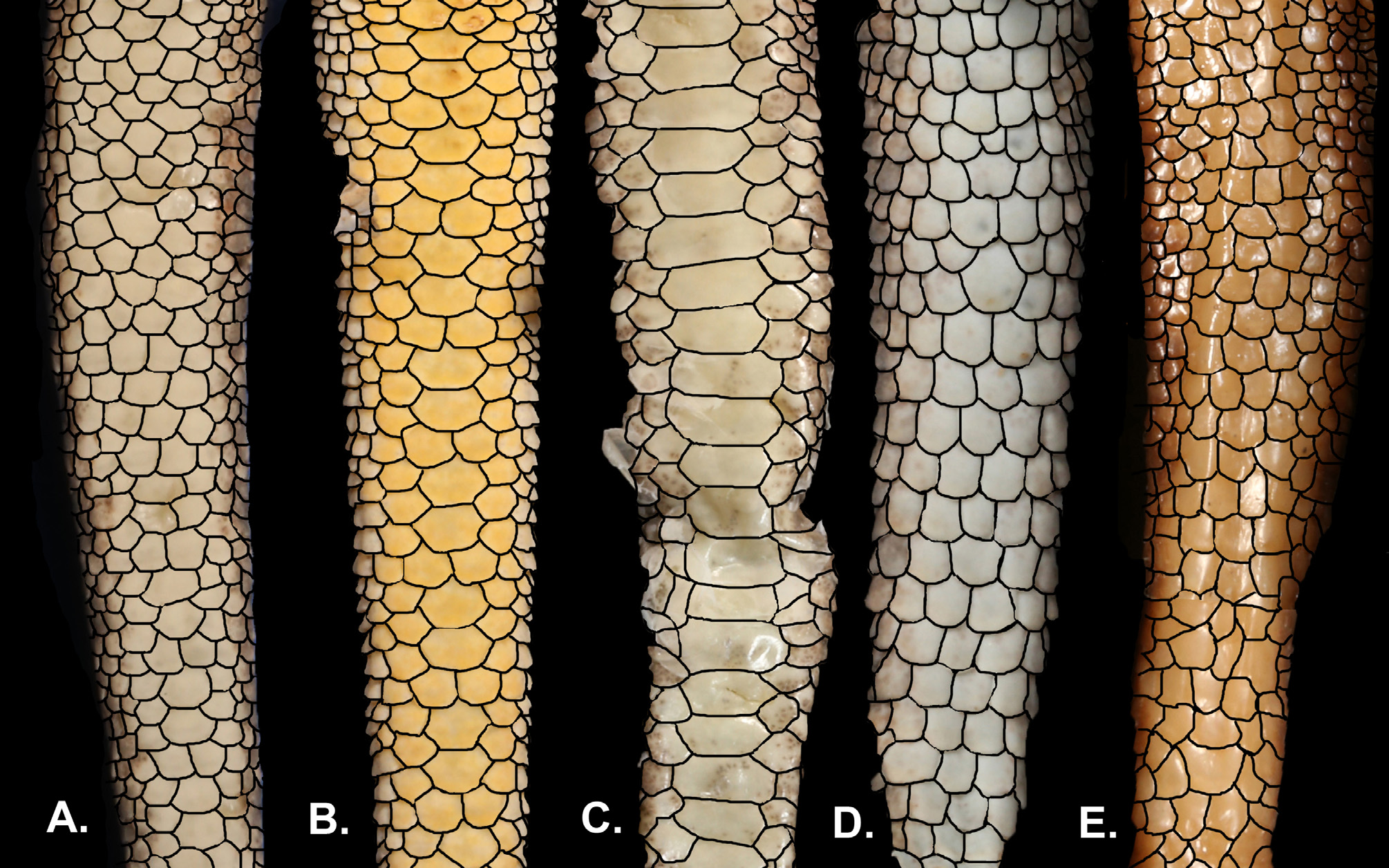
Diagnosis. Lygodactylus nyaneka sp. nov. is a small dwarf gecko with a maximum SVL of 34 mm. It can be distinguished from other members of the genus occurring in Angola and surroundings region by having 1) four scales surrounding the nostril (1 st supralabial, two supranasal and one postnasals); 2) nostrils separated from the rostral and pierced above the suture between rostral and first supralabial; 3) mental followed by three postmentals; 4) first infralabial with 25–50% overlap of postpostmental; 5) 16 to 17 rows of ventral scales at midbody; 6) 4–6 precloacal pores in males; 7) median subcaudals arranged in a pattern of one row of scales about ¼ width of the tail alternating with two rows of scales about ½ width of the tail ( Fig. 4B View FIGURE 4 ); 8) venter white; 9) gular region usually white, but sometimes speckled with minute black dots. Background usually grey to brown with a pair of black lines on the flanks bordering a pale stripe (not always visible) with a line of cream dots from the posterior part of the orbit to the end of the tail ( Figs 14–15 View FIGURE 14 View FIGURE 15 ).
Comparison with other South and Southwestern African Lygodactylus . As the molecular data provide evidence of the distinctiveness of Lygodactylus nyaneka sp. from all other taxa, we here restrict our morphological comparisons to those named congeners occurring in Angola and surrounding countries. Lygodactylus nyaneka sp. nov. differs from L. angolensis , L. chobiensis , L. lawrencei and L. tchokwe sp. nov. by having 4–6 precloacal pores ( versus 7 – 10 in L. angolensi s; 7 – 11 in L. chobiensis ; two in L. lawrencei ; two to four in L. tchokwe sp. nov.) ( Fig. 2 View FIGURE 2 ). The new species can be distinguished from L. baptistai sp. nov., L. bradfieldi and L. capensis in not having the rostral in contact with the nostrils. It can be further distinguished from nominotypical L. capensis by its staggered rows of subcaudal plates in original tails ( versus a more linear distribution of the rows in L. capensis ). It differs from L. chobiensis and L. baptistai sp. nov. by its white ventral and gular coloration ( versus bright yellow in L. chobiensis and L. baptistai sp. nov.).
Description of holotype. SVL 34.1 mm; TrunkL: 16.7 mm; CrusL 5.3 mm; TL: 27.2 mm; TW: 3.1 mm; HL: 8.4 mm; HW: 5.7 mm; HH: 3.8 mm; EarL: 0.5 mm; ForeaL: 4.5 mm; ED: 1.8 mm; NE: 2.5 mm; SE: 3.9 mm; EE: 2.6 mm; IN: 1.1 mm; IO: 3.1 mm. Head broad and slightly distinct from neck; snout longer than broad, the distance from its edge to the anterior border of the eye greater than the interorbital distance anteriorly, and greater than the distance between the eye and ear opening; ear opening medium and oval; snout covered with enlarged homogeneous flattened granular scales, becoming smaller laterally above the eye; scales on the snout slightly larger than those on the occiput; prominent canthus rostralis; rostral pentagonal, wider than high; four scales surrounding the nostril (1 st supralabial, two supranasal and one postnasals); nostrils separated from the rostral and pierced above the suture between rostral and first supralabial ( Fig. 3 View FIGURE 3 ); mental pentagonal, longer than broad, followed by three postmentals; scales behind postmentals intermediate in size, decreasing towards base of the throat and increasing on chest and belly; supralabials 8; infralabials 7; first infralabial with 25% to 50% contact with postpostmental; gulars smooth, enlarged and forming transverse rows posteriorly, 26 between posterior extent of the infralabials/jaws; 16 rows of ventral scales at midbody; limbs short but well developed, extremities pentadactyl; 1 st digit of both manus and pes extremely reduced and lacking claw, distal portions of remaining digits expanded, bearing claw; 5 pair of lamellae under 4 th of digit manus and 5 pair of lamellae beneath 4 th of digit pes; dorsum with small, homogeneous, granular and keeled scales with a similar size to those on trunk, the scales on limbs are slightly larger; venter with enlarged, smooth, homogeneous scales; scales on ventral surface of the fore- and hindlimbs are smaller to those on venter, as well as the surface scales of thighs that are slightly smaller and imbricate; precloacal pores 6 ( Fig. 2 View FIGURE 2 ); approximately 18-19 enlarged generation glands present along the ventral surface of each thigh; tail cylindrical; posterior half regenerated; dorsal caudal scales enlarged, flattened and homogeneous; keeled, spinose and acuminate on the lateral surface of the tail; median subcaudals scales flattened and smooth, one row of scales about ¼ width of tail alternating with two rows of scales about ½ width of tail.
Coloration in ethanol. The coloration of Lygodactylus nyaneka sp. nov. is brown to ashy-grey above, with a pair of black lines on the flanks from snout to the end of the tail, bordering a pale stripe with a line of cream dots that continues to the tail. Mid-dorsal region with small scattered cream dots. The head slightly darker than body and limbs, bearing a speckled pattern with some lighter irregular dots. The venter is white with no markings or speckling, although lateral margins of the belly and ventral surface of the limbs are speckled. The chin and gular regions white with no markings or speckling. Dorsal surface of the limbs with scattered cream dots. Tail brown with a pattern of two parallel cream dots, running on across the dorsal surface up to the regenerated part, ventrally white with no markings or speckling.
Variation. Variation in scalation and body measurements of the type series is reported in Table 6. Paratypes agree almost entirely with the holotype, with the exception of two male paratypes ( CAS 263529 About CAS ; UF 187228) that differ slightly in coloration, with both specimens having a darker dorsum with a more faded pattern, and speckled black on the gular region ( CAS 263529 About CAS ) .
Distribution. In Angola the species is known from the central and southwestern regions of the country, from the lower areas of Benguela to Namibe, Huíla and Bié provinces, and potentially to Cuando Cubango Province. Based on geography, the single specimen identified by Baptista et al. (2019) from Bicuar National Park main camp as Lygodactylus angolensis most likely represent a misidentification of Lygodactylus nyaneka sp. nov. In neighboring Namibia, the species is known with certainty from Epupa Falls ( Fig. 16 View FIGURE 16 ) but is probably more extensively distributed.
Habitat and natural history notes. The species is usually found basking during daytime on rocky outcrops, trees, logs or human infrastructures, such as fences, houses and poles ( Fig. 17 View FIGURE 17 ). Never found on the ground. This species occurs in sympatry with Lygodactylus angolensis , in the northern region of Bié Province.
Etymology. The specific epithet “ nyaneka ” is a noun in apposition and is given in honor of the dominant ethnolinguistic group of the region where the species exists. We suggest “Nyaneka Dwarf Gecko” and “Osga-Anã-Nhanheca” as the English and Portuguese common names, respectively, for this species.



| AMB |
Asenovgrad Museum |
| T |
Tavera, Department of Geology and Geophysics |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lygodactylus nyaneka
| Marques, Mariana P., Ceríaco, Luis M. P., Buehler, Matthew D., Bandei- Ra, Suzana A., Janota, Joyce M. & Bauer, Aaron M. 2020 |
Lygodactylus angolensis ( Baptista et al. 2019: 113 )
| Baptista, N. L. & Antonio, T. & Branch, W. R. 2019: 113 |
Lygodactylus capensis bradfieldi ( FitzSimons 1943: 53 )
| FitzSimons, V. F. 1943: 53 |
Lygodactylus capensis capensis
| Bates, M. F. & Branch, W. R. & Bauer, A. M. & Burger, M. & Marais, J. & Alexander, G. J. & de Villiers, M. S. 2014: 118 |
| Schmidt, K. P. 1933: 4 |
Lygodactylus capensis
| Marques, M. P. & Ceriaco, L. M. P. & Blackburn, D. C. & Bauer, A. M. 2018: 190 |
| Branch, W. R. 1998: 246 |
| Frade, F. 1963: 253 |
| Loveridge, A. 1957: 187 |
| Loveridge, A. 1947: 208 |
| Monard, A. 1937: 53 |
| Bocage, J. V. B. 1895: 15 |
Hemidactylus capensis
| Bocage, J. V. B. 1870: 68 |
| Bocage, J. V. B. 1867: 219 |