
Protoptilinae Ross, 1956: 149
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3723.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:27DD0C15-FFEA-4079-88F7-ADD726A5E98F |
|
persistent identifier |
https://treatment.plazi.org/id/197E87B9-CA72-FF9D-6FAB-59E2FD777428 |
|
treatment provided by |
Felipe |
|
scientific name |
Protoptilinae Ross, 1956: 149 |
| status |
|
Subfamily Protoptilinae Ross, 1956: 149 View in CoL
The monophyly of Protoptilinae has been corroborated by this study. Monophyly was recovered in all analyses of all datasets, and strongly supported in all except the parsimony analysis of the SUBSET COI dataset. This study corroborated 2 of the 5 putative synapomorphies of Protoptilinae identified by Morse & Yang (1993) including: 1) foretibial spur hair-like or absent ( Fig. 6B View FIGURE 6 ) (character 17), 2) forewing crossveins forming a relatively straight line along anastomosis ( Fig. 7A View FIGURE 7 ) (character 47). Morse & Yang (1993) also listed an absence of apical fork V in the forewing as a possible synapomorphy of Protoptilinae , but this analysis did not support that character. Additionally, since larval characters were not included in this study, we were unable to evaluate the putative larval characters (tarsal claw seta beside process; anal claw with 4 accessory hooks) suggested by Morse & Yang (1993). Another unique synapomorphy of Protoptilinae identified in this study is the presence of a row of erect setae along the Cu 2 in the forewing ( Fig. 7A View FIGURE 7 ) (character 41).
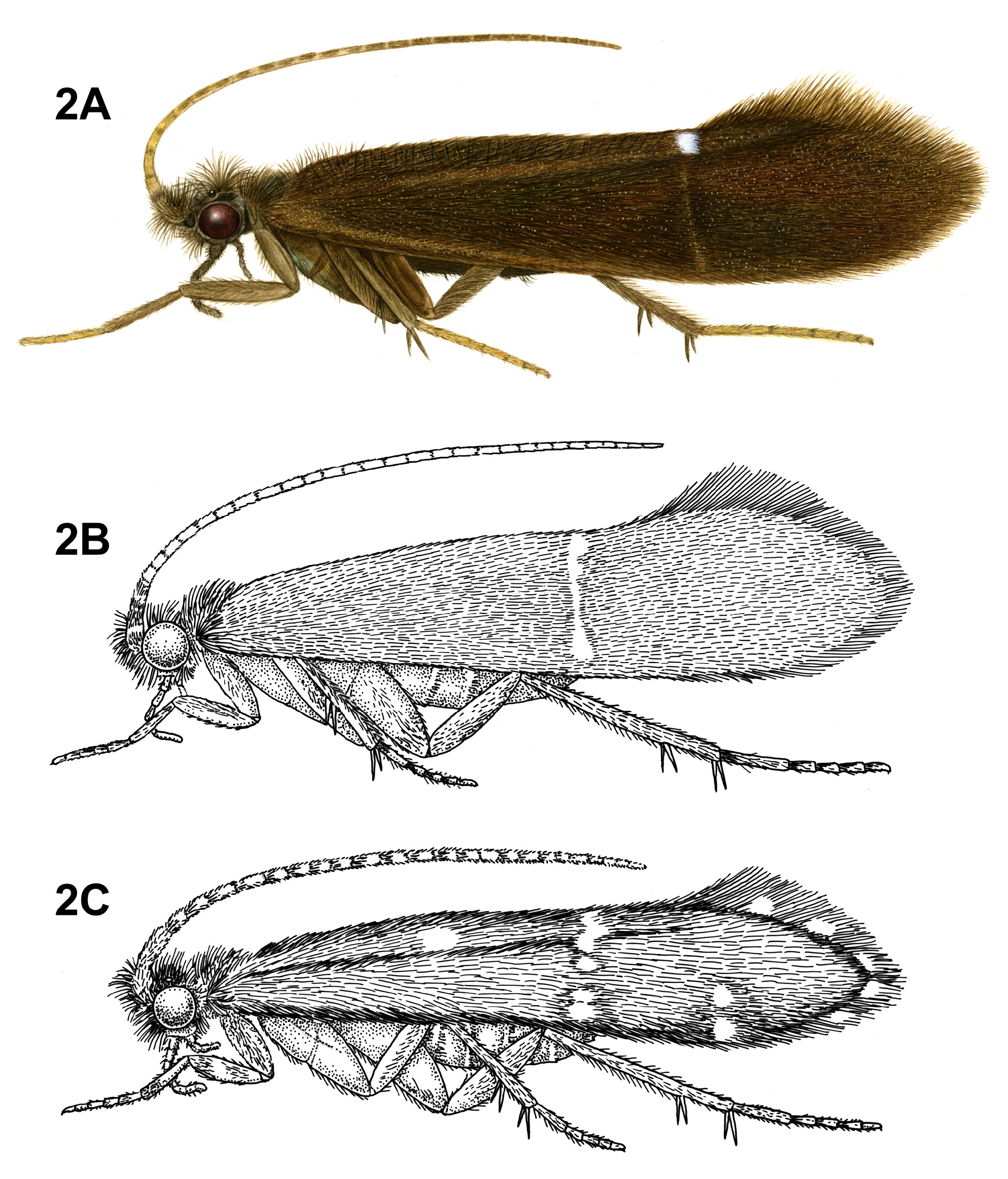
Diagnosis of Protoptilinae . Protoptilines are very minute caddisflies. As such, they may be confused with members of the family Hydroptilidae ; however, protoptilines can easily be separated based on their presence of mesocutal setal warts. Perhaps the most identifiable feature of Protoptilinae is the absence of a foretibial spur, or a hair-like condition of this spur. Other distinctive features include the row of erect setae along Cu 2 in the forewing, and the linear aspect of the forewing crossveins.
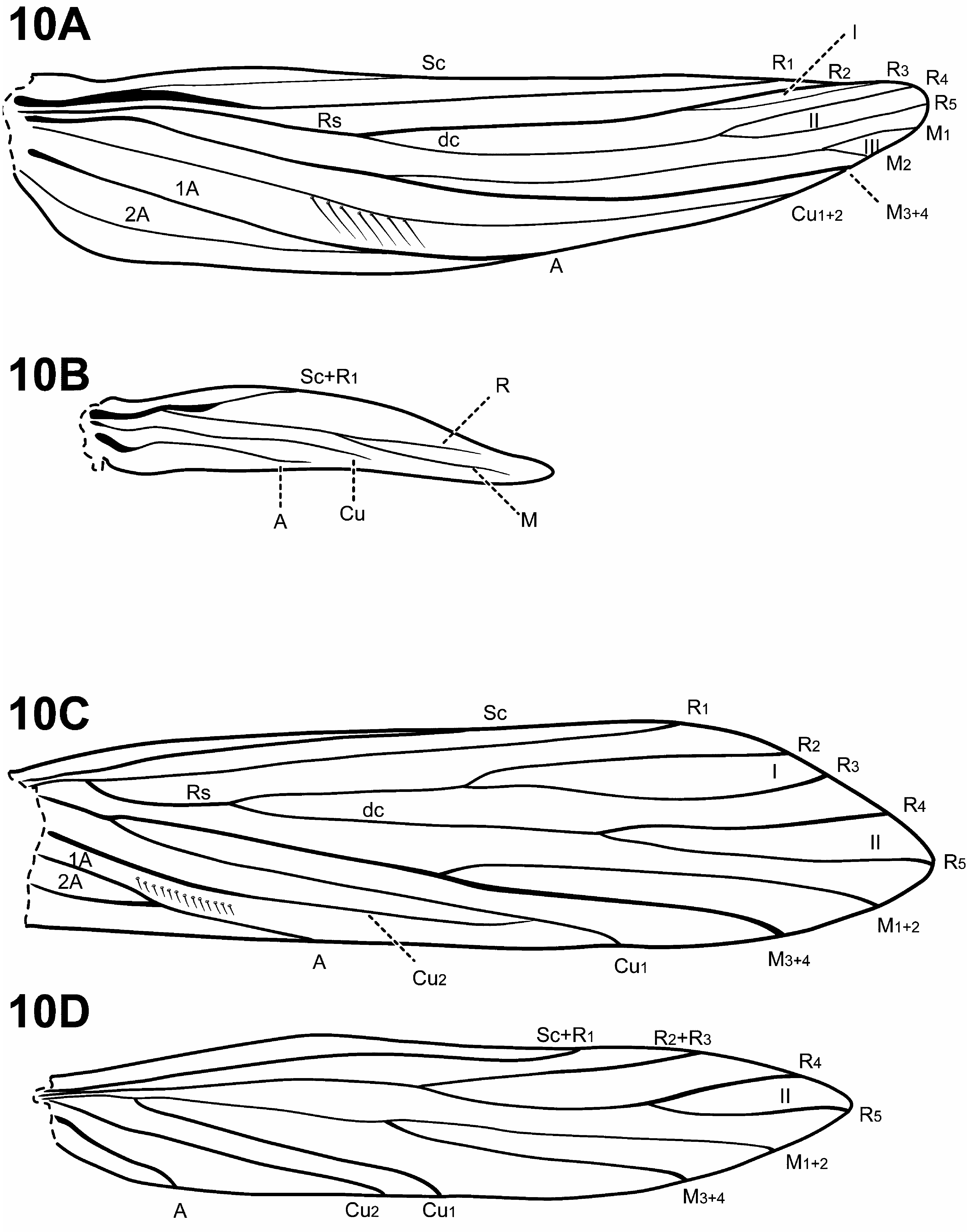
Adult. Length of forewing: 1.5– 6 mm. Body, wings, and appendages pale brown, tawny brown, or fuscous, often intermingled with rufous or golden hairs. Wings often with few pale cream-colored or white hairs, specks or spots, and transverse line along anastomosis. Forewing with long fringe of setae along apical margin; hind wing with long fringe of setae along posterior margin. Head broader than long, vertex rounded. 3 ocelli. Mesal setal warts of pronotum widely spaced, not touching mesally. Mesoscutum with 2 pairs of setal warts; mesoscutellar setal warts usually absent, or if present, small and round. Maxillary palps 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Tibial spur formula variable; foretibial preapical spur absent, foretibial apical spur either absent or hair-like. Fore- and hind wing venation variable, ranging from complete to extremely reduced. Forewing R1 unforked; crossveins, when visible, forming relatively straight line along anastomosis; row of stout, erect setae present along Cu2. Male genitalia extremely variable among genera and species. Preanal appendages absent.
Genus Canoptila Mosely, 1939
Canoptila Mosely, 1939: 218 View in CoL [Type species: Canoptila bifida Mosely, 1939 View in CoL , by original designation].
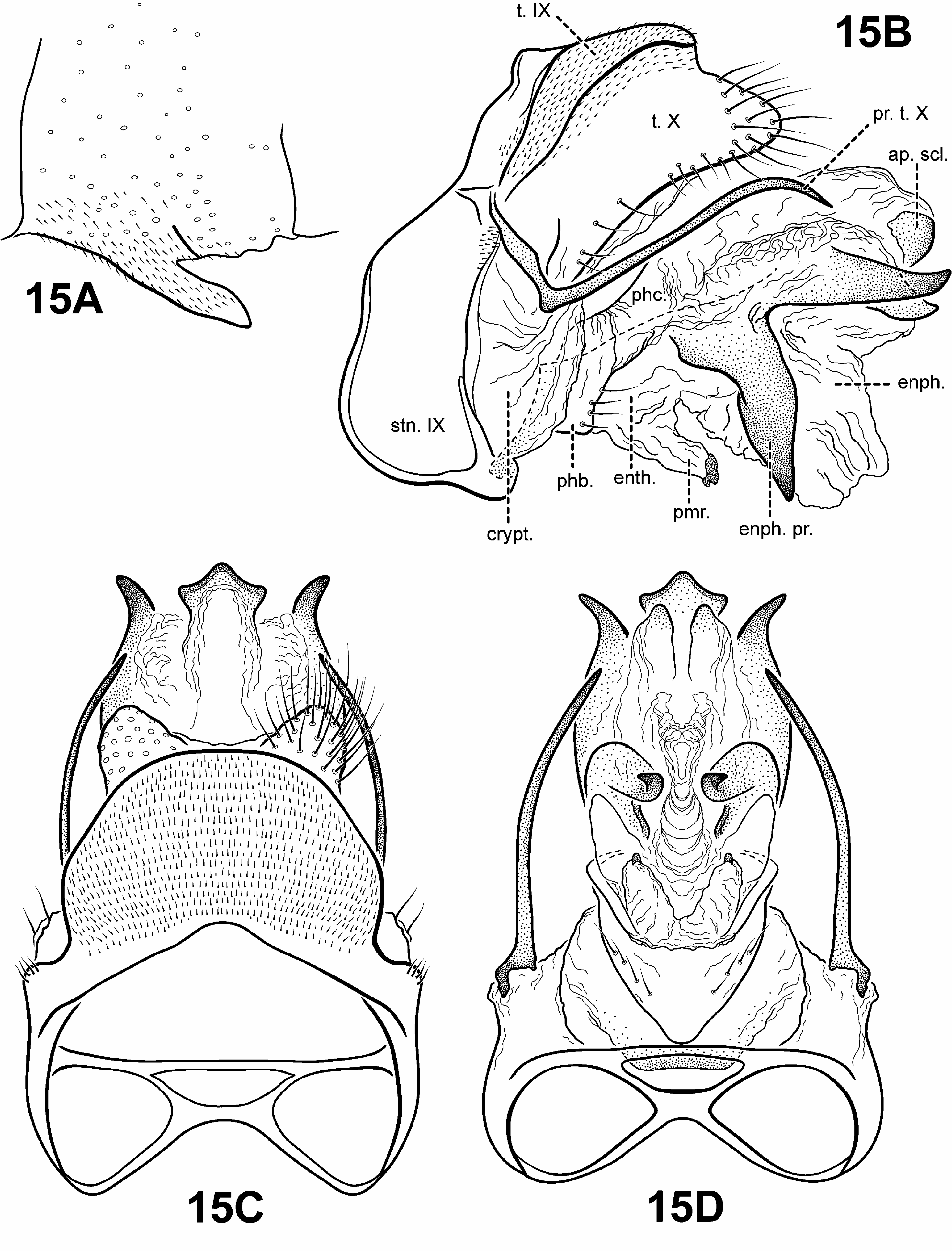
All of the analyses recovered a strongly supported monophyletic Canoptila View in CoL . A weak sister relationship between Scotiotrichia View in CoL and Canoptila View in CoL was recovered in the parsimony analysis of the TOTAL MORPH dataset. The analysis confirmed a putative unique synapomorphy identified by Robertson & Holzenthal (2006): the presence of membranous and digitate parameres (character 86) ( Figs. 15B, D View FIGURE 15 ). However, the presence of long spine-like posterolateral process on tergum X and wing venational features were not found to be unambiguous synapomorphies of Canoptila View in CoL .
Diagnosis of Canoptila ( Figs. 5B View FIGURE 5 , 9A, 9B View FIGURE 9 , 15 View FIGURE 15 , 31B View FIGURE 31 ). As noted by Robertson and Holzenthal (2006), the genus Canoptila is easily identified by the presence of certain structures of the male genitalia ( Fig. 15 View FIGURE 15 ): 1) a pair of long spine-like processes on the posterolateral margins of tergum X, and 2) paired, highly membranous digitate parameres arising basoventrally on the endotheca. Canoptila is most similar in forewing venation ( Fig. 9A View FIGURE 9 ) to Cariboptila , Itauara , and Mastigoptia based on the presence of forks I, II, and III, the intersection of Cu1 and Cu2 near anastomosis, and absence of A3. However, Canoptila can be distinguished from these species by the length of the stem of fork II: in Canoptila the stem is longer than the fork whereas in the other genera, the stem is shorter or no longer than the fork. Hind wing ( Fig. 9B View FIGURE 9 ) venation of Canoptila is nearly identical to that of Scotiotrichia , and is very similar to those of Cariboptila and Protoptila having only apical fork II present and only 1 anal vein. Although indistinguishable from Scotiotrichia , the hind wings of Cariboptila and Protoptila are narrow and scalloped past the anastomosis, whereas in Canoptila , the margins are nearly parallel. Canoptila has telescopic glandular structures ( Fig. 5B View FIGURE 5 ) arising from the tegulae, as in many species of Culoptila . However, in Culoptila , these structures are “concertina-shaped” ( Mosely 1954) and positioned posterolaterally, whereas in Canoptila , they are tubular in shape and arise more anteriorly.
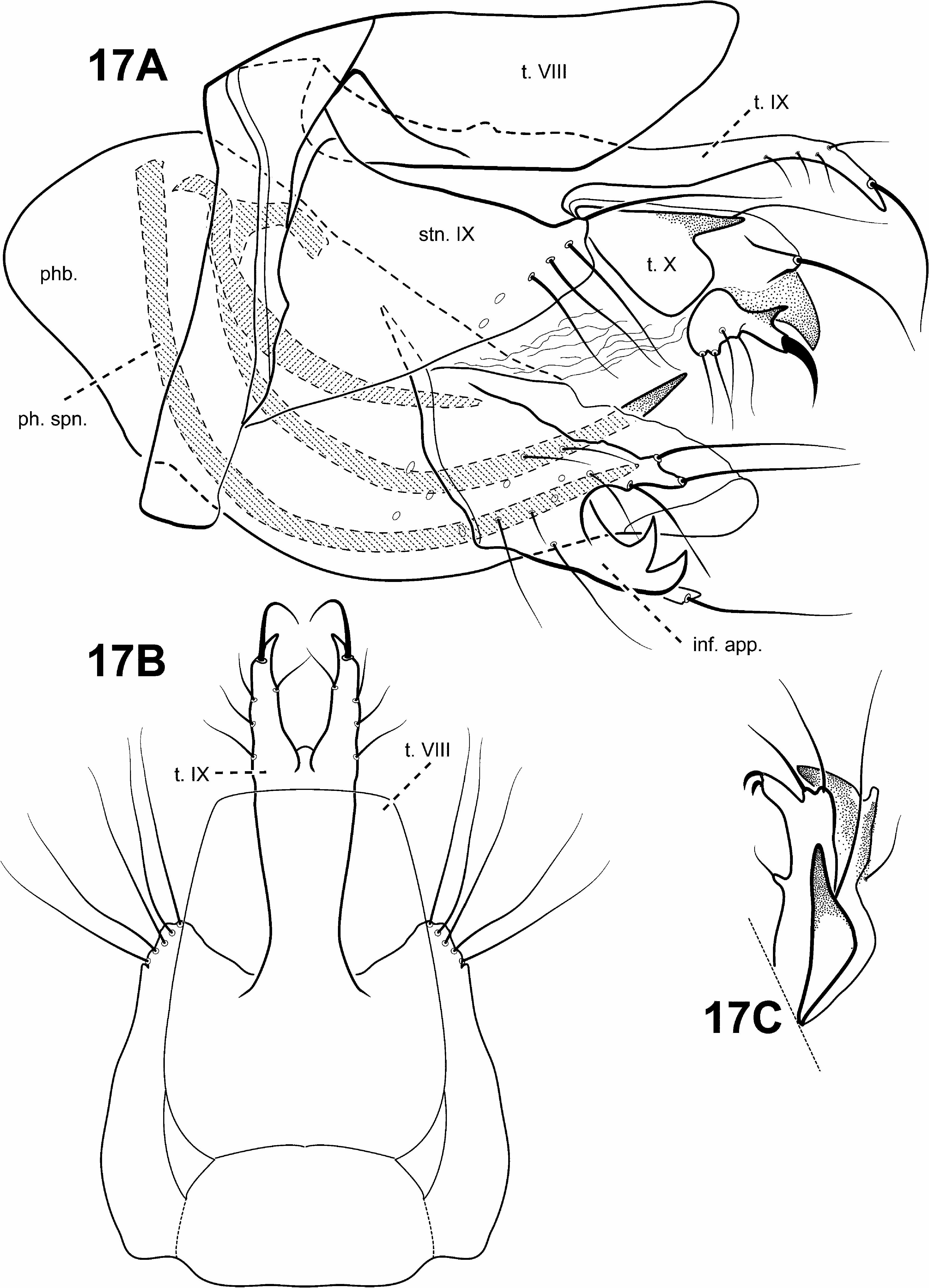
Adult. Body, wings, and appendages nearly uniformly fuscous or tawny brown, tibia and tarsi yellowish brown. Head ( Fig. 5B View FIGURE 5 ) broader than long, vertex rounded, with large anteromesal setal wart, 2 distinct pairs of suboval anterior setal warts, small suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5- segmented, 1st and 2nd segments short with elongate setae apically; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts. Mesothorax ( Fig. 5B View FIGURE 5 ) wider than long, with paired telescopic tegular glands; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 9A View FIGURE 9 ) relatively narrow, with margins nearly parallel, apex oblique. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate, stem longer than fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present below Cu2; A3 absent; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 9B View FIGURE 9 ) margins nearly parallel, tapering only slightly past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1,4,4, foretibial spur extremely reduced and hairlike. Sternum VI process ( Fig. 15A View FIGURE 15 ) elongate and digitate, apex subacute, associated with weak oblique apodeme posteriorly.
Male genitalia ( Fig. 15 View FIGURE 15 ). Preanal and inferior appendages absent. Segment IX anterior margin fairly straight; tergum IX well developed, relatively broad, simple, without processes; sternum IX uniformly narrow, mesally, without modification. Tergum X completely fused to tergum IX forming ridge at line of fusion; dorsomesal margin bifid or subquadrate; dorsolateral margin with highly setose, rounded or subquadrate lobes; ventrolateral margin with paired very elongate, spine-like process directed inwardly. Parameres present, arising laterally from endotheca, membranous, bulbous, with sclerotized or asperous apices. Phallobase reduced, lightly sclerotized, with paired row or patches of setae ventrolaterally. Endophallus highly membranous, enlarged and convoluted when evaginated, bearing 1 to several pairs of large, pointed, sclerotized processes, lightly sclerotized apically or with terminal sclerite.
Female genitalia ( Fig. 31B View FIGURE 31 ). (Female known only for C. williami .) Truncate posteriorly, not extensible. Abdominal segment VIII short, synscleritous, posterolateral margin slightly incised. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Cariboptila Flint, 1964
Cariboptila Flint, 1964: 17 View in CoL [Type species: Cariboptila orophila Flint, 1964 View in CoL , by original designation]
Campsiophora Flint, 1964: 14 View in CoL [Type species: Campsiophora pedophila Flint, 1964 View in CoL , by original designation] new synonym
Cubanoptila Sykora, 1973 View in CoL , in Botosaneanu & Sykora 1973: 383 [Type species: Cubanoptila cubana Sykora, 1973 View in CoL , by new, subsequent designation] new synonym
Muangpaipsyche Malicky & Silalom, 2012: 22 [Type species: Muangpaipsyche areopagita Malicky & Silalom, 2012 , by monotypy; Malicky 2013, as synonym of Campsiophora View in CoL ] new synonym
None of the analyses of any dataset in this study were able to recover a monophyletic Cariboptila as historically constituted. Flint (1964) established Cariboptila and Campsiophora in the same paper, and distinguished the 2 genera based on 1) the level of branching of the apical forks 1 and 2 relative to each other in the forewing, and 2) the presence of a hair pencil on the inner surface of the forewing in Campsiophora . However, differences in the relative position of the apical forks are quite variable and homoplastic throughout the subfamily and 1 of the 2 states must be plesiomorphic and thus not appropriate for defining taxa by cladistic standards. Additionally, the hair pencil in the forewing is not consistently found in all Campsiophora . Sykora (in, Botosaneanu & Sykora 1973), distinguished Cubanoptila from other protoptiline genera based on features of the antennae, wing venation, and a plate-like tergum VIII. While the presence of small spines on the 3rd antennal article does appear to be unique to Cubanoptila , the other characters proposed for the establishment of Cubanoptila are not and, in these analyses, the Cubanoptila species are nested within Cariboptila , and thus the groups are not reciprocally monophyletic at the generic level. Furthermore, larval morphology of the 3 genera is not significantly different, with all 3 possessing tarsal claws with a short and broad seta. Therefore, to reflect phylogeny accurately, the following species are hereby transferred to Cariboptila (all new combinations): from Campsiophora : C. arawak (Flint) , C. mulata (Botosaneanu) , C. pedophila (Flint) ; and from Cubanoptila : C. botosaneanui (Kumanski) , C. cubana (Sykora) , C. madremia (Botosaneanu) , C. muybonita (Sykora) , C. purpurea (Sykora) , C. tridens (Botosaneanu) . Although simultaneously published, as first revisers we prefer the name Cariboptila as the senior synonm over Campsiophora , despite “page priority.” The use of Cariboptila will impart more nomenclatural stability since fewer species need to be transferred. Furthermore, the name alludes to their occurrence in the Caribbean.
Malicky & Silalom (2012) described a new monotypic genus, reportedly from Thailand, Muanpaipsyche areopagita . However, after we informed Dr. Malicky that this new genus is almost certainly Cariboptila pedophila ( Flint, 1964) because of nearly identical male genitalia and wing venation (compare figures given by Malicky & Silamon 2012, page 23 with our Figs. 8 View FIGURE 8 & 18 View FIGURE 18 ), Malicky synonomized the genus, but not the species ( Malicky 2013). We hereby designate the species C. arepagita , as a new junior synonom of C. pedophila . It is extremely doubtful that this single species would be found both in the Caribbean and Thailand, but is rather the likely result of a mislabeled specimen or other curatorial lapse.
No type species was designated explicitly in the original description of Cubanoptila . Flint et al. (1999) erred in stating that Cubanoptila cubana is the type species by “original designation.” Cubanoptila cubana is here designated as the type species of Cubanoptila by “virtual tautonomy” in compliance with Recommendation 69A.2 of the International Code of Zoological Nomenclature (1999).
Monophyly of the redefined Cariboptila was strongly supported in all analyses with the following 3 unique synapomorphies: 1) antennal scape greater than or equal to 3 times the length of the pedicel ( Fig. 6A View FIGURE 6 ) (character 3), 2) tergum IX dorsal lateral process bearing 1 or more elongate apical setae ( Figs. 16A, C View FIGURE 16 ) (character 73), 3) inferior appendages present as broad, highly setose, prominent plate-like projections fused basoventrally to phallobase ( Figs. 16A, B View FIGURE 16 ) (character 82).
Diagnosis of Cariboptila ( Figs. 6A View FIGURE 6 , 7C, 7D View FIGURE 7 , 8 View FIGURE 8 , 16–18 View FIGURE 16 View FIGURE 17 View FIGURE 18 ). Perhaps the most distinctive feature of the genus Cariboptila is its extremely long antennal scape ( Fig. 6A View FIGURE 6 ), which is often associated with androconia. Many species also have stout setae along the 3 rd antennal segment, although this is not completely diagnostic for the genus. Another diagnostic feature is the short discoidal cell in the forewing. The males of 2 species of Padunia [ P. falcata (Schmid, 1991) and P. phyllis (Malicky & Chantaramongkol, 2007) ] also have a short discoidal cell, but in these species the wings are highly modified. Cariboptila is most similar in forewing venation to Canoptila , Itauara , and Mastigoptia based on the presence of forks I, II, and III, the intersection of Cu1 and Cu2 near the anastomosis, and the absence of A3. However, Cariboptila can be distinguished from these genera based on its presence of a short discoidal cell in the forewing. The hind wing venation of Cariboptila is nearly identical to that of Canoptila , Scotiotrichia , and Protoptila , having only apical fork II and only a single anal vein, yet can be differentiated from these genera based on differences of the forewing. In the forewing of Protoptila , A3 is present; in Scotiotichia, fork III is absent; and Canoptila ’s forewing has a short discoidal cell.
Diagnostic characters of the male genitalia include the broad, highly setose plate-like inferior appendages, the digitate process of tergum IX, which usually bears 1 or more elongate apical setae, and the lateral branches of tergum X whose dorsolateral margins have sclerotized short, flattened, inwardly curved processes or irregular setose processes and ventrolateral margins bearing 1 or several large, highly sclerotized spines or spine-like setae. The male genitalia of Cariboptila are most similar to those of Culoptila in that both are completely lacking, or have a strap-shaped, sternum IX, and an enlarged phallobase. The 2 genera are easily differentiated by differences in the shapes of terga IX and X, the inferior appendages, and wing venation. The genera Tolhuaca and Scotiotrichia also have a reduced sternum IX and enlarged phallobase, but can be easily separated from Cariboptila by other genitalic features or differences in wing venation. Tolhuaca has complete forewing venation and the phallic apparatus lacks elongate spines or processes. In the forewing of Scotiotrichia , fork III is absent.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs. Wings often with few pale cream-colored or white hairs, spots, or transverse line along anastomosis and small white specks or spots along apical margin. Head broader than long, vertex rounded, either with 1 distinct pair, 1 divided pair, or 2 distinct pairs of suboval anterior setal warts, small or large suboval posterior warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape more than 3 times as long as pedicel, often with androconia; 3rd antennal segment occasionally with stout spine-like setae ( Fig. 6A View FIGURE 6 ). Maxillary palps 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large, subtriangular or suboval pronotal setal warts. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval, or occasionally elongate anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Figs. 7C View FIGURE 7 , 8A, 8C View FIGURE 8 ) relatively narrow, with margins nearly parallel, apex oblique. Males occasionally with hair pencil along anal margin ( Fig. 8C View FIGURE 8 ) or callosity in anal or apical costal region of forewing. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile or only slightly petiolate with extremely short stem; fork II petiolate or sessile, but when petiolate, stem shorter or about as long as fork; fork III petiolate, stem usually longer than fork, occasionally same length; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis, or completely fused; row of erect setae present along Cu2; A3 absent; crossveins forming relatively linear transverse cord; discoidal cell shorter than Rs vein. Hind wing ( Figs. 7D View FIGURE 7 , 8B, 8D View FIGURE 8 ) narrow and scalloped past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs either 0,4,4 or 1,4,4, foretibial spur extremely reduced and hair-like. Sternum VI process present, shape variable, ranging from flattened dorsoventrally, short and digitate, to thumb-like and prominent, apex also variable, either pointed, rounded, or subquadrate, often associated with weak oblique apodeme posteriorly.
Male genitalia ( Figs. 16–18 View FIGURE 16 View FIGURE 17 View FIGURE 18 ). Preanal appendages absent. Tergum VIII occasionally forming dorsal plate subtending tergum IX and X ( Fig. 17A, 17B View FIGURE 17 ). Sternum VIII without modification. Segment IX anterior margin rounded or fairly straight, posterolateral margin without lateral process or lobes; tergum IX well developed, relatively broad, with paired, dorsolateral, often digitate process usually bearing 1 or more elongate apical setae; sternum IX usually completely absent, or if present, consisting only of small ventral membranous strap. Tergum X subtended by tergum IX; dorsomesal margin bifid or subquadrate, or with single broad, plate-like process, or irregular with several small processes; dorsolateral margin with sclerotized short, flattened, inwardly curved process, or with irregular setose processes; ventrolateral margin bearing 1 or several large, highly sclerotized spines or spine-like setae, directed mesad. Inferior appendages present, ventrally as broad, highly setose, prominent platelike projection fused ventrobasally to phallobase, often invaginated apicomesally, with setose, elongate, broad and plate-like, or multilobed lateral appendages. Parameres absent. Phallobase extremely enlarged, sclerotized, and occupying nearly all of genital capsule, forming ventral portion of genitalia. Phallic apparatus with 1 to several pairs of elongate phallic spines and/or highly curving processes. Endophallus membranous, enlarged and convoluted when invaginated.
Female genitalia. (Females unknown for many species.) Truncate posteriorly, not extensible. Abdominal segment VIII broad or short, sometimes incomplete midventrally with elongate sclerite. Sternum IX often forming triangular lobes. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Culoptila Mosely, 1954
Culoptila Mosely, 1954: 336 View in CoL [Type species: Culoptila aluca Mosely, 1954 View in CoL , by original designation].
Monophyly for Culoptila View in CoL was strongly supported in all analyses with the following unique synapomorphies: 1) an incomplete Cu1 on the forewing (character 38), and 2) inferior appendages comprised of simple, paired, long or short process, fused to one another basally and to ventral surface of phallobase (character 82). Blahnik and Holzenthal (2006) hypothesized that Culoptila View in CoL was most closely related to the Caribbean genera Campsiophora View in CoL , Cariboptila View in CoL , and Cubanoptila View in CoL . In this study, a sister relationship between Culoptila View in CoL and the Caribbean taxa was recovered with weak nodal support in the parsimony analysis of the TOTAL MORPH dataset, and in both analyses of the SUBSET MORPH dataset.
Diagnosis of Culoptila ( Figs. 3B, 3C View FIGURE 3 , 4 View FIGURE 4 , 7A 7B View FIGURE 7 , 19 View FIGURE 19 ). The genus Culoptila can be identified by both forewing and hind wing venation. In the forewing, only apical forks I–IV are present, fork V is absent. Additionally, Cu1 is incomplete, not reaching the wing margin. In the hind wing, only apical forks II and III are present. Another distinctive feature, although not completely diagnostic for the genus as already noted by Blahnik and Holzenthal (2006), is the presence of enlarged tegulae on the mesothorax of males, which accommodate an extensible concertina-like glandular structure. The genus Canoptila also possesses extensible glandular structures associated with the tegulae, but these differ in shape from those of Culoptila .
Other diagnostic features of the genus occur in the male genitalia, most notably the absence or extreme small size of sternum IX, and a greatly enlarged phallobase. As noted above, and in the works by Robertson & Holzenthal (2005) and Blahnik & Holzenthal (2006), the genera Cariboptila and Tolhuaca have similar reductions of the sterna and inflated phallobases; however, these genera can be easily separated from Culoptila based on differences in wing venation and other characters of the male genitalia. Terga IX and X are differently shaped in Cariboptila , and unlike Culoptila , Cariboptila lacks apical fork III in the hind wing. Tolhuaca has complete forewing venation and lacks elongate phallic spines. Another diagnostic feature of Culoptila is the presence of an apical phallotremal sclerite, although this structure is sometimes difficult to see and may not be easily identified ( Blahnik & Holzenthal 2006).
Adult. Body, wings, and appendages pale brown to fuscous. Wings often with white transverse line along anastomosis or conspicuous spot at the arculus. Head broader than long, vertex rounded, with pair of small anteromesal setal warts, pair of distinct, suboval anterior setal warts, small suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts. Mesothorax shape variable, usually wider than long, but occasionally longer than wide to accommodate enlarged tegulae of males bearing paired concertina-shaped glandular processes; mesoscutum with pair of elongate anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 7A View FIGURE 7 ) relatively narrow, with margins nearly parallel, apex oblique. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I, II, III, and IV present; Sc and R1 distinct along their entire lengths; fork I petiolate, but with extremely short stem; fork II petiolate or sessile, stem shorter than fork; fork III petiolate, stem longer than fork; fork IV petiolate, stem about as long as fork; Cu1 incomplete, not reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 7B View FIGURE 7 ) narrow and scalloped past anastomosis; apical forks II and III present; Sc and R1 fused basally; A2 present, not looped. Tibial spurs 1,4,4, foretibial spur extremely reduced and hair-like. Sternum VI process present, short, often somewhat circular in lateral view, apex subquadrate or rounded, associated with weak oblique apodeme posteriorly. Male genitalia ( Fig. 19 View FIGURE 19 ). Preanal appendages absent. Segment IX anterior margin rounded; tergum IX well developed, relatively broad, simple, without processes; sternum IX absent. Tergum X incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin subquadrate; dorsolateral margin without processes; ventrolateral margin with paired elongate processes attached ventrolaterally to tergum IX and directed ventrad and sometimes anterad. Inferior appendages present (except in C. cantha Ross, 1938 , and C. plummerensis Blahnik & Holzenthal, 2006 ), as simple, paired long or short appendages, fused to one another basally and to ventral surface of phallobase. Parameres absent. Phallobase extremely enlarged, lightly sclerotized, and occupying nearly all of genital capsule, with sclerotized, projecting apex posterodorsally. Phallic apparatus with 1 or 2 phallic spines of varying length and shape.
Female genitalia. (Females unknown for many species.) Truncate posteriorly, not extensible. Abdominal segment VIII short, usually synscleritous, but sometimes incomplete midventrally and laterally. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Itauara Müller, 1888
Itauara Müller, 1888: 275 View in CoL [Type species: Antoptila brasiliana Mosely, 1939 View in CoL , subsequent selection by Flint et al. 1999]. Antoptila Mosely, 1939: 219 View in CoL [Type species: Antoptila brasiliana Mosely, 1939 View in CoL , by original designation] Flint et al. 1999, to synonymy.
A monophyletic group consisting of 21 Itauara species was recovered in all analyses of this study. The unique synapomorphy of Itauara View in CoL is the presence of a dorsal sheath covering a ventral membranous portion of the phallus (character 95).
Diagnosis of Itauara ( Figs. 9C, 9D View FIGURE 9 , 20 View FIGURE 20 ). The genus Itauara can be identified by features of the male genitalia. The phallic apparatus consists of a sclerotized dorsal sheath covering a very membranous ventral portion, an apparent posterior extension of the phallobase or phallicata. In some species, this sclerotized dorsal sheath seems to detach from the ventral membrane apically to reveal a single dorsomesal process or spine (e. g., I. amazonica Flint, 1971 ). Mortoniella has a similar dorsomesal process or spine, but in Mortoniella it arises internally from the phallobase, whereas in Itauara it appears to arise dorsobasally from this sclerotized sheath. In some species ( I. guarani and I. plaumanni ), the sheath produces a dorsolateral flange, although this character is not diagnostic for the genus. Another genitalic feature characteristic of Itauara is an extremely reduced phallobase. In most species, the phallobase is barely visible, consisting of a small, very lightly sclerotized, or entirely membranous structure. The genera Mastigoptila and Canoptila display similar reductions or absences of the phallobase, but can easily be separated from Itauara by other genitalic characters: Mastigoptila has an elongate, whip-like process arising from the membranes of the phallocrypt; Canoptila has highly membranous digitate parameres. When present (they are absent in many species), the inferior appendages are rather distinct for Itauara , consisting of a single or apically bifid process produced mesally and fused to the phallobase basoventrally.
The forewing venation of Itauara is most similar to that of Cariboptila and Canoptila , with apical forks I–III and a lack of 3A. Canoptila can be differentiated from Itauara by having stout setae occurring below Cu2 whereas in Itauara the setae occur along the vein. Cariboptila can be differentiated from Itauara by the presence of a short discoidal cell, that of Itauara being long. The hind wing venation of Itauara is variable, with either apical forks II, III, and V or II and V or III only ( I. amazonica ) or II only.
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi yellowish brown ( Fig. 2A View FIGURE 2 ). Wings usually with partial white transverse line along anastomosis not reaching costal margin, or with conspicuous white spot at arculus ( Fig. 2A View FIGURE 2 ). Head broader than long, vertex rounded, with pair of small anteromesal setal warts or with large anteromesal setal wart, either 1 distinct pair or 1 divided pair of suboval anterior setal warts, small or large suboval posterior warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 9C View FIGURE 9 ) usually relatively narrow, with margins nearly parallel, occasionally narrowed past anastomosis or much reduced, apex oblique or rounded. Male occasionally with callosity present in apical costal region of forewing. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile or only slightly petiolate with extremely short stem; fork II petiolate or sessile, but when petiolate, stem length variable; fork III petiolate, stem variable in length; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present along Cu2; A3 absent; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 9D View FIGURE 9 ) margins nearly parallel, tapering only slightly past anastomosis, or narrow and scalloped past anastomosis; venation variable, with either apical forks II, III, and V present or II and V present or III present ( I. amazonica ) or II present; Sc and R1 fused basally or converging near wing margin; A2 absent. Tibial spurs 1,4,4, rarely 1,3,4, foretibial spur extremely reduced and hair-like. Sternum VI process present, short and digitate or thumb-like and prominent, apex rounded or attenuate and pointed, usually associated with oblique apodeme posteriorly.
Male genitalia ( Fig. 20 View FIGURE 20 ). Segment IX anterior margin rounded, posterolateral margin without lateral processes or lobes; tergum IX usually not well developed, simple, and without processes; sternum IX without modification, except in I. brasiliana ( Mosely, 1939) , which bears 2 pairs of elongate, seta-like processes. Tergum X incompletely fused to tergum IX ventrolaterally or rarely ( I. amazonica ) completely fused and indistinguishable from tergum IX, shape extremely variable; dorsomesal margin may be simple without processes, bifid apicomesally with single broad plate-like process, or irregular with several small processes; dorsolateral margin either simple structure without processes, or more commonly with small paired lobes, elongate and down-turned and finger-like processes, or irregular setose processes; ventrolateral margin with paired elongate or broad flange-like processes directed ventrad and sometimes anterad, or with 1 or more irregular, paired, setose, digitate lobes directed posterad. Inferior appendages either present or absent; when present, consisting of single or apically bifid process produced mesally, broadest at base and fused to phallobase ventrobasally. Parameres present except in I. brasiliana , arising either ventrobasally from phallobase or laterally from endotheca, sclerotized, shape variable. Phallobase extremely small and difficult to discern. Phallic apparatus with sclerotized dorsal sheath covering membranous ventral portion, sometimes receding to a single dorsomesal process arising dorsobasally from phallobase, phallicata occasionally with dorsolateral flange, or occasionally with dorsomesal spine arising posterior of phallobase. Endophallus highly membranous, enlarged and convoluted when evaginated, occasionally bearing small apical spine-like sclerites and processes.
Female genitalia. (Females unknown for many species.) Truncate posteriorly, not extensible. Abdominal segment VIII short, synscleritous, posterolateral margin slightly incised. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Mastigoptila Flint, 1967
Mastigoptila Flint, 1967: 49 [Type species: Mastigoptila curvicornuta Flint, 1967 , by original designation].
Monophyly of Mastigoptila was supported in all analyses in this study. A synapomorphy of Mastigoptila is markedly asymmetrical inferior appendages, fused to the phallocrypt ventrobasally (character 82) ( Fig. 20A View FIGURE 20 ).
Diagnosis of Mastigoptila ( Figs. 12C, 12D View FIGURE 12 , 21 View FIGURE 21 ). The most distinctive feature of Mastigoptila is the profoundly asymmetrical aspect of the male genitalia. The phallic apparatus is tubular, asymmetrical, and often arcuate. The inferior appendages form an asymmetrical complex consisting of differently shaped and sized right and left setose appendages, which are fused together basally and ventrobasally with a lightly sclerotized phallocrypt. In some species, 1 of the inferior appendages also bears an elongate spinelike process. Another identifying feature is the presence of an elongate whiplike process arising from membranes of the phallocrypt on 1 side of the genitalia. The phallobase is also apparently absent.
The forewing venation of Mastigoptila is similar to that of Canoptila and identical to that of some species of Itauara . The forewing of Mastigoptila can be differentiated from that of Canoptila by having a sessile apical fork II. Unlike Mastigoptila , Itauara has symmetrical genitalia.
Adult. Body, wings, and appendages nearly uniformly fuscous. Wings sometimes with light spot at arculus and faint transverse line along anastomosis. Head broader than long, vertex rounded, pair of distinct, suboval anterior setal warts, small suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5-segmented, 1st and 2nd segments short with elongate setae apically; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts. Mesothorax wider than long, of some species with tegular glands; mesoscutum usually with pair of elongate anteromesal setal warts, although occasionally entire anteromesal region setose with no distinct patch, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing relatively narrow, with margins nearly parallel, apex oblique. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II sessile; fork III petiolate, stem about as long as fork, occasionally slightly longer; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present below Cu2; A3 absent; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing margins nearly parallel, tapering only slightly past anastomosis; apical forks II, III, and V present; Sc and R1 converging near wing margin; A2 absent. Tibial spurs either 0,4,4 or 1,4,4; foretibial spur extremely reduced and hair-like. Sternum VI process present, flattened laterally, apex attenuate and pointed, often associated with weak oblique apodeme posteriorly. Sternum VII occasionally with small mesal point.
Male genitalia. Preanal appendages absent. Segment IX anterior margin rounded, posterolateral margin without lateral process or lobes; tergum IX strap-like, simple, without processes; sternum IX uniformly narrow, mesally without modification. Tergum X incompletely fused to tergum IX, with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin subquadrate or excavate; dorsolateral margin either hood-like, without processes or with 1 or more large, horn-like processes; ventrolateral margin usually simple, but occasionally bearing small lobes or processes. Inferior appendages present, markedly asymmetrical, consisting of elongate spine-like processes and setose lobes, sometimes bifid, fused to phallocrypt ventrobasally. Parameres absent. Phallobase apparently absent. Phallic apparatus asymmetrical, tubular and often arcuate with posteriorly projecting apex, sclerotized or rugose, with small membranous protuberances, highly convoluted internal membranes with occasional small spines.
Female genitalia. (Females unknown for most species.) Truncate posteriorly, not extensible. Abdominal segments VIII and IX not fused.
Genus Merionoptila Schmid, 1959
Merionoptila Schmid, 1959: 482 View in CoL [Type species: Merionoptila wygodzinskyi Schmid, 1959 View in CoL , by original designation].
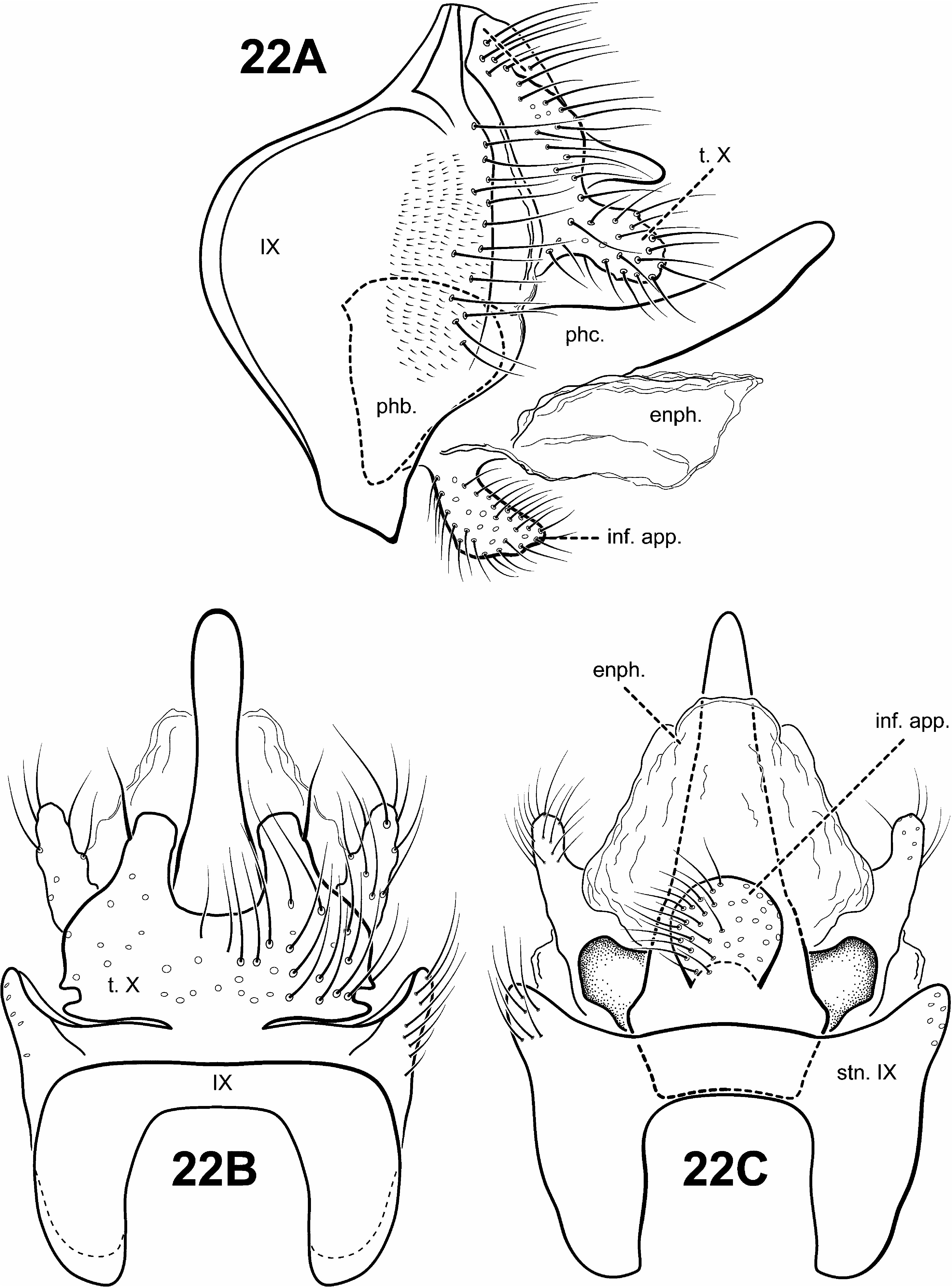
Schmid (1959) suggested that the monotypic genus Merionoptila View in CoL was more closely related to Antoptila View in CoL (= Itauara View in CoL ) than to the other known protoptiline genera. This study can neither refute nor support that possibility since the placement of Merionoptila View in CoL remains unresolved. The following are apomorphies of Merionoptila View in CoL : 1) stem of forewing fork I longer than fork (character 26) ( Fig. 10A View FIGURE 10 ), 2) forewing crossvein r-m absent (character 45) ( Fig. 10A View FIGURE 10 ), 3) sternum VI mesal process absent, and 4) inferior appendage bulbous, fused ventromedially to the endotheca and projecting ventrad (character 82) ( Fig. 22A, C View FIGURE 22 ).
Diagnosis of Merionoptila ( Figs. 10A, 10B View FIGURE 10 , 22 View FIGURE 22 ). This monotypic genus has quite distinctive morphology among protoptilines. Its thorax is quite broad, it has highly setose legs and eyes, and extremely reduced wings—all apparently adaptations for skating across the water surface, a behavior first reported by Wygodzinski in a letter to Schmid (1959). The forewing is reduced in size, but has rather typical venation, with apical forks I–III present, although fork I may be difficult to see. The hind wing is even more reduced, and it is nearly impossible to discern the venation. The male genitalia of Merionoptila are rather simple, with a sclerotized, tubular phallic apparatus, directed dorsally at the apex. Inferior appendages are quite distinct, consisting of a highly setose, bulblike process, projecting ventrad and fused basally to the ventral portion of the phallobase.
Adult. Body, wings, and appendages nearly uniformly fuscous. Head broader than long, vertex rounded, with pair of small anteromesal setal warts, pair of distinct, suboval anterior setal warts, small suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Eyes setose. Maxillary palps 5-segmented, 1st membranous and bulbous, 2nd segment very short; last 3 segments each nearly same length as 1st and 2nd segments combined. Thorax broad and robust. Prothorax with 2 large, subtriangular or suboval pronotal setal warts. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 10A View FIGURE 10 ) much reduced, apex rounded. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I, II, and III present, fork I very difficult to discern; Sc and R1 distinct along their entire lengths; fork I petiolate; fork II petiolate, stem longer than fork; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 completely fused; row of erect setae present along Cu2; A3 absent; crossveins difficult to discern, but apparently forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 10B View FIGURE 10 ) extremely reduced; apical forks not clearly identifiable; Sc and R1 fused basally; A2 absent. Mesothoracic legs extremely setose. Tibial spurs 0,2,4. Sternum VI process absent.
Male genitalia ( Fig. 22 View FIGURE 22 ). Preanal appendages absent. Segment IX anterior margin rounded; tergum IX uniformly narrow, simple, without processes; sternum IX uniformly narrow, mesally without modification. Tergum X fused to tergum IX dorsomesally, or "hinged"; dorsomesal margin divided or bifid apicomesally; dorsolateral margin with small paired lobes; ventrolateral margin with irregular, paired setose lobes. Inferior appendages present, bulbous, highly setose, fused ventromedially to endotheca and projecting ventrad. Parameres absent. Phallobase simple, subtriangular in lateral view. Phallic apparatus simple, tubular, projecting posterad, without spines or processes. Endophallus membranous, enlarged and convoluted when invaginated.
Female genitalia. (A female specimen for this monotypic genus has not been positively associated with the male. In the original description, Schmid reported that a single female was captured, but at a different locality from the type specimens captured. Thus, only basic female genitalic characters are briefly described.) Truncate posteriorly, not extensible.
Genu s Mortoniella Ulmer, 1906
Mortoniella Ulmer, 1906: 95 View in CoL [Type species: Motoniella bilineata , by monotypy].
Mexitrichia Mosely, 1937: 158 View in CoL [Type species: Mexitrichia leroda View in CoL , by original designation] Blahnik & Holzenthal 2008, to synonymy.
Paraprotoptila Jacquemart, 1963: 342 [Type species: Paraprotoptila armata , by monotypy] Flint et al. 1999, to synonymy with Mexitrichia View in CoL .
A monophyletic Mortoniella including a single undescribed species from Brazil was recovered with weak support in analyses of the TOTAL COMBO and SUBSET COMBO datasets. However, in analyses of the TOTAL MORPH and SUBSET MORPH datasets, Mortoniella was found to be paraphyletic, and in the SUBSET COI dataset, polyphyletic. Blahnik and Holzenthal (2008) synonomized Mexitrichia with Mortoniella based on similarities of the male genitalia and the fact that Mexitrichia was historically defined based on the presence of apical fork V in the hind wing, a plesiomorphic character for Trichoptera . It is perhaps the retention of this and other primitive characters (e. g., an unmodified tergum X and sternum VIII), that accounts for the difficulty in recovering its monophyly in some datasets. We sampled species from both Mortoniella and Mexitrichia , as previously defined. Based on morphological evidence, and the fact that a monophyletic Mortoniella that included the Mexitrichia species was recovered in combined analyses, the decision by Blahnik and Holzenthal (2008) to synonymize Mexitrichia with Mortoniella is justified by this study. A synapomorphy of Mortoniella is that the inferior appendages form a composite structure consisting of paired processes fused together basally and to the ventral margin of the phallic apparatus and ventrolaterally to the endotheca, with associated articulated appendages fitting into pockets (character 82) ( Figs. 23A, D View FIGURE 23 ).
Diagnosis of Mortoniella ( Figs. 2B View FIGURE 2 , 11A, 11B View FIGURE 11 , 23 View FIGURE 23 ). The genus Mortoniella is diagnosed based on several unique structures of the male genitalia, termed the “phallic ensemble” by Blahnik and Holzenthal (2008). The male genitalia are characterized by the presence of a dorsomesal spine or process arising internally from the phallobase, which varies in shape among species. The dorsomesal spine or process articulates with the phallicata, which sometimes bears a dorsolateral process that may function as a guide for the spine ( Blahnik & Holzenthal 2008). Tergum X is usually excavated dorsomesally to accommodate the spine. Some species of Itauara have a similar dorsomesal spine, but in this genus, the spine arises posteriorly as an apparent extension of the phallobase. The inferior appendages of Mortoniella are also distinct among protoptilines. They are fused to one another basally and to the ventral part of endotheca, and enclose a pair of sclerotized pockets on the mesal surface. These pockets are associated with pair of small, digitate, articulated appendages arising from the posteroventral part of the phallobase. Members of the genus Protoptila also bear the small articulated appendages that fit into associated pockets. However, in Protoptila , the inferior appendages are apparently absent. The 2 genera can also be separated based on differences in the shape of tergum X, the lack of a dorsomesal spine in Protoptila , and an unmodified sternum VIII, which is posteriorly projecting in Protoptila .
The forewing venation of Mortoniella is most similar to that of Protoptila , with apical forks I, II, and III present, a completely fused Cu1 and Cu2, and the presence of 3A (although in some species of both genera, 3A is absent). However, Mortoniella can sometimes be distinguished by the row of erect setae, which is positioned only slightly below the Cu2 vein in Mortoniella , but far below the Cu2 vein in Protoptila . The hind wing venation of Mortoniella is quite variable, with either apical forks II, III, and V or II and III only or II only present.
Adult. Body, wings, and appendages nearly uniformly fuscous or tawny brown. Wings often with white transverse line along entire length of anastomosis ( Fig. 2B View FIGURE 2 ). Head broader than long, vertex rounded, either 1 distinct pair or 1 divided pair of suboval anterior setal warts, large suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 11A View FIGURE 11 ) shape variable, relatively broad past anastomosis in most species, more narrow in others, apex rounded, occasionally with scale-like setae. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II sessile; fork III petiolate, stem variable in length; Cu1 complete, reaching wing margin; Cu1 and Cu2 completely fused; row of erect setae present slightly below Cu2; A3 when present, looped; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 11B View FIGURE 11 ) shape variable, relatively broad past anastomosis in most species, more narrow with margins nearly parallel in other species; venation variable, either with apical forks II only, or II and III only, or II, III, and V present; Sc and R1 fused basally; A2 when present, looped. Tibial spurs either 0,4,4 or 1,4,4; foretibial spur extremely reduced and hair-like. Sternum VI process present, shape variable, subtriangular, elongate and digitate, short and digitate, or laterally flattened, apex rounded or attenuate and pointed, usually associated with oblique apodeme posteriorly.
Male genitalia ( Fig. 23 View FIGURE 23 ). Preanal appendages absent. Segment IX anterior margin rounded, posterolateral margin without lateral process or lobes; tergum IX uniformly narrow, simple, without processes; sternum IX uniformly narrow, mesally without modification. Tergum X fused to tergum IX dorsomesally, or "hinged"; dorsomesal margin excavated, divided or bifid apicomesally, but sometimes with single, prominent, elongate process; dorsolateral margin without processes, as small paired lobes, or with irregular setose processes; ventrolateral margin without processes, with irregular, paired setose lobes, or with 1 or more irregular, paired, setose, digitate lobes directed posterad. Inferior appendages present, forming composite structure consisting of paired processes fused together basally, ventrolaterally to endotheca, and to ventral margin of phallobase associated with articulated appendages. Phallobase with small pair of articulated digitate, rod-like appendages with membranous apices, associated with modified pockets ventrobasally. Parameres present, arising dorsolaterally from endotheca, sclerotized, sinuous or mostly straight. Phallic ensemble with single dorsomesal spine or process emerging internally from phallobase. Endophallus membranous, enlarged and convoluted when invaginated, sometimes with sclerotized regions and spines.
Female genitalia. (Females unknown for many species.) Truncate posteriorly, not extensible. Abdominal segment VIII short, synscleritous, posterolateral margin slightly incised, dorsal and ventral margins sometimes invaginated posteromesally. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Padunia Martynov, 1910
Padunia Martynov, 1910: 425 View in CoL [Type species: Padunia adelungi Martynov, 1910 View in CoL , by monotypy]
Matrioptila Ross, 1956: 164 View in CoL [Type species: Protoptila jeanae Ross, 1938 View in CoL , by original designation] new synonym
Nepaloptila Kimmins, 1964: 37 View in CoL [Type species: Nepaloptila coei Kimmins, 1954 View in CoL , by original designation] new synonym
Poeciloptila Schmid, 1991: 243 View in CoL [Type species: Poeciloptila falcata Schmid, 1991 View in CoL , by original designation] new synonym
Temburongpsyche Malicky, 1995: 15 View in CoL [Type species: Temburongpsyche anakan Malicky, 1995 View in CoL , by original designation] new synonym
Uenotrichia Tsuda, 1942: 228 [Type species: Uenotrichia fasciata Tsuda, 1942 , by monotypy]; Fischer, 1971, as senior synonym of Padunia View in CoL ; Marshall, 1979, as junior synonym of Padunia View in CoL .
The Asian Clade is comprised of genera from the East Palaearctic and Oriental regions ( Padunia View in CoL , Nepaloptila View in CoL , Poeciloptila View in CoL , Temburongpsyche View in CoL ) and the Nearctic region ( Matrioptila View in CoL ). In all the TOTAL datasets, a paraphyletic Nepaloptila View in CoL is basal to a subclade containing these remaining genera. Within this subclade, Padunia View in CoL as traditionally constituted, was recovered as a paraphyletic taxon in all analyses of the TOTAL datasets. Additionally, a sister relationship was consistently found between the type species, Padunia adelungi View in CoL , and Matrioptila jeanae View in CoL . Relationships of the remaining 5 Padunia species included in the analyses were either unresolved or paraphyletic, interspersed with species of Poeciloptila View in CoL and Temburongpsyche View in CoL .
Kimmins (1964) remarked that the male genitalia of Nepaloptila resembled those of Matrioptila but differentiated the 2 genera based on Nepaloptila ’s retention of fork V in the forewing. However, complete wing venation is plesiomorphic for Trichoptera and thus this character is not appropriate for defining Nepaloptila by current cladistic standards.
Likewise, Ross (1956) defined Matrioptila based on several plesiomorphic characters (retention of fork V in the hind wing, separate course of Cu 2 in forewing, and distinct claspers in male genitalia). Although Matrioptila can be distinguished from the other Asian genera by the absence of fork V in the forewing, this fork apparently has been lost independently several times in the evolution of Protoptilinae lineages and, at this taxonomic level, may simply be autapomorphic for the species.
Poeciloptila was primarily defined by 2 male genitalic features: 1) membranous lateral faces of segment X, which enables segment X to “rock downwards,” and 2) a large ventral branch of phallus (Schmid 1991). However, examination of Poeciloptila revealed the lateral faces of tergum X to be lightly sclerotized and immobile, rather than membranous, a characteristic that is not uncommon in other protoptiline genera. Furthermore, the ventral branch of the phallus is not unique to Poeciloptila , being present in some species of Padunia , Nepaloptila , and Matrioptila .
Malicky (1995) established Temburonpsyche as a new genus based on its broad, ring-like segment IX, reduction of various “appendices” in the form of “finger”-like processes along the margin of segment IX, a large phallus, and tibial spur formula 0,3,3. Yet, a broad ring-like segment IX is plesiomorphic in Trichoptera . Additionally, many of these genera have a somewhat enlarged phallus and the various finger-like processes along the margin of segment IX are most likely homologous to similar processes in some species of the other genera, which have also apparently undergone a reduction. Finally, Matrioptila shares the 0,3,3 spur formula, so this character is not unique to Temburongpsyche . Furthermore, Malicky (1995) also remarked that the wing venation of Temburongpsyche corresponds closely to that of Padunia and Poeciloptila , and the head and thoracic warts are similar to those of Nepaloptila .
To sum, many of these genera were defined solely on plesiomorphic characters or features that are not unique to the particular taxon. Although the larvae of Nepaloptila , Poeciloptila and Temburongsyche are unknown, the larvae of Matrioptila and Padunia are very similar—both construct dorsoventrally flattened cases, have “trifid” tarsal claws with 3 equally sized processes, and similarly shaped mesonotal sclerites. Furthermore, the female genitalia of these 5 genera are very similar in shape. Therefore, to reflect phylogeny accurately, the following species are hereby transferred to Padunia (all new combinations): from Matrioptila : P. jeanae (Ross) , from Nepaloptila : P. coei (Kimmins) , P. jisunted (Malicky & Chantaramongkol) , P. kanikar (Malicky & Chantaramongkol) , P. ruangjod (Malicky & Chantaramongkol) ; from Poeciloptila : P. almodad (Malicky & Chantaramongkol) , P. atyalpa (Schmid) , P. eringena (Malicky & Silalom) , P. falcata (Schmid) , P. maculata (Tian & Li) , and P. phyllis (Malicky & Chantaramongkol) ; and from Temburongpsyche : P. anakan (Malicky) . Additionally, Padunia briatec (Malicky & Chantaramongkol) , which was transferred to Poeciloptila by Schmid (1991), is hereby returned to Padunia (restored combination).
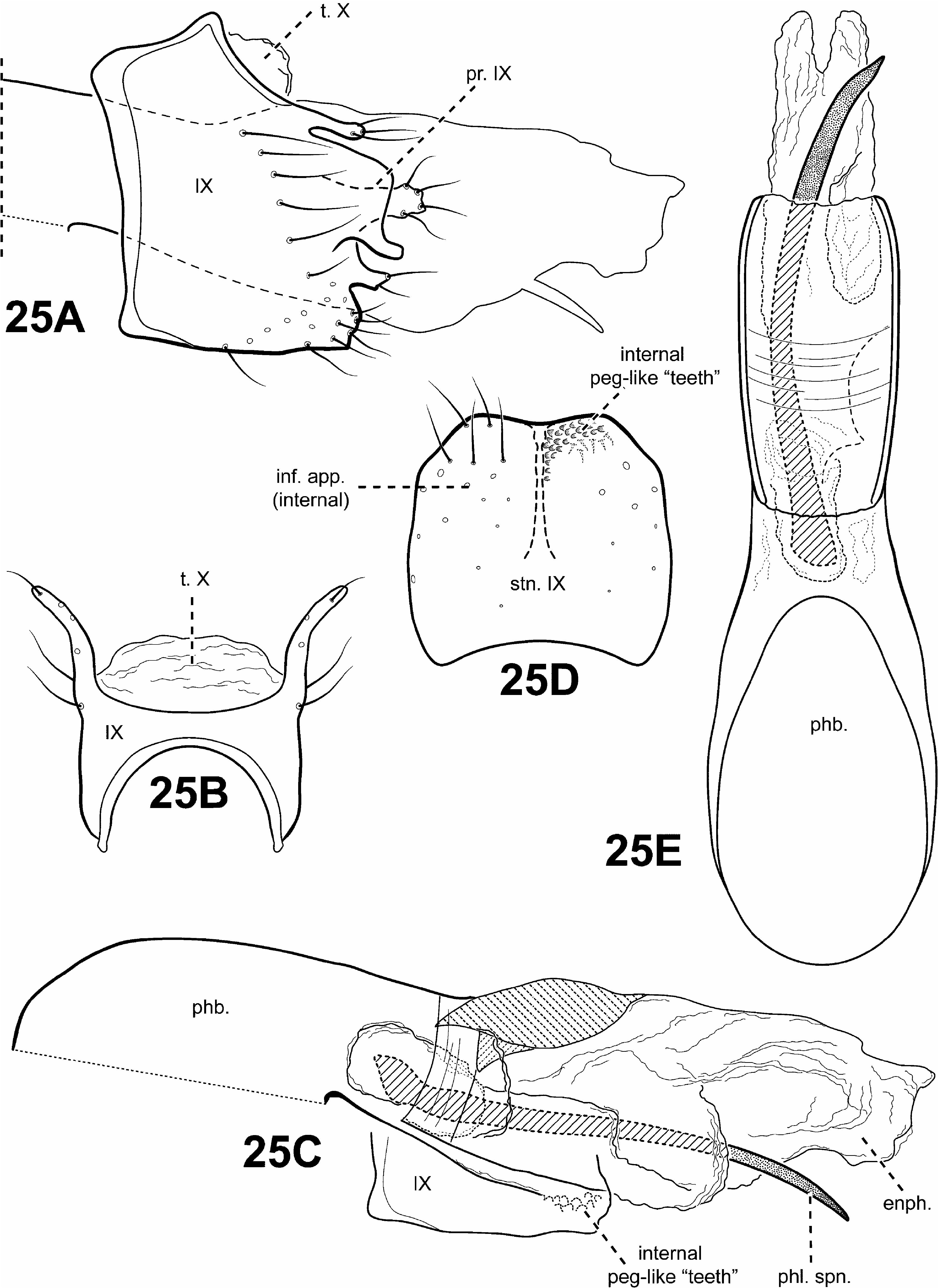
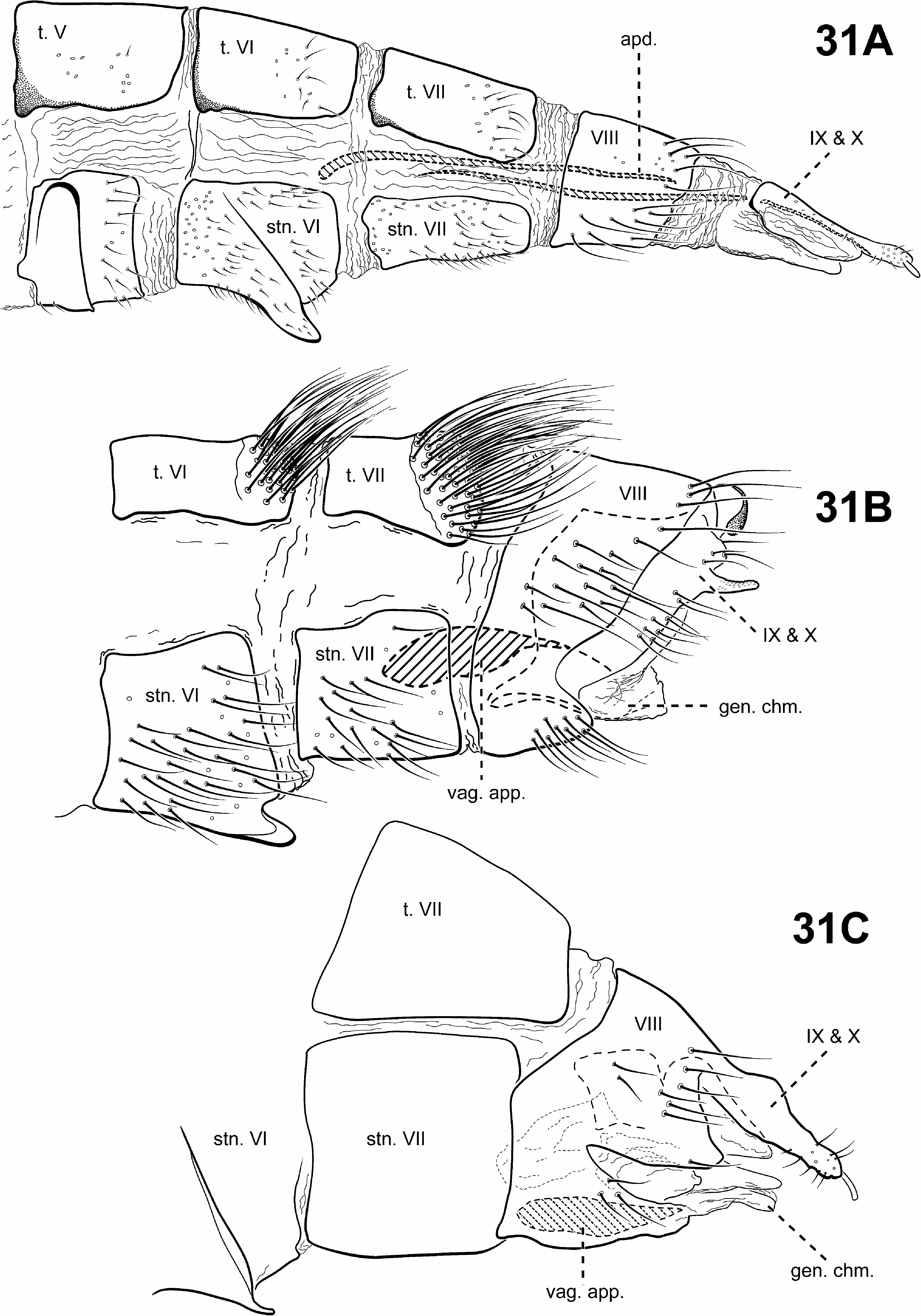
Monophyly of this newly defined Padunia was recovered in all analyses based on the following synapomorphies: 1) the phallobase has its dorsal margin forming a sclerotized sheath that is produced anteriorly, with its ventral margin membranous or absent (character 90) ( Figs. 24A, B View FIGURE 24 ); 2) segments VIII and IX of the female genitalia are fused (character 99) ( Fig. 31C View FIGURE 31 ).
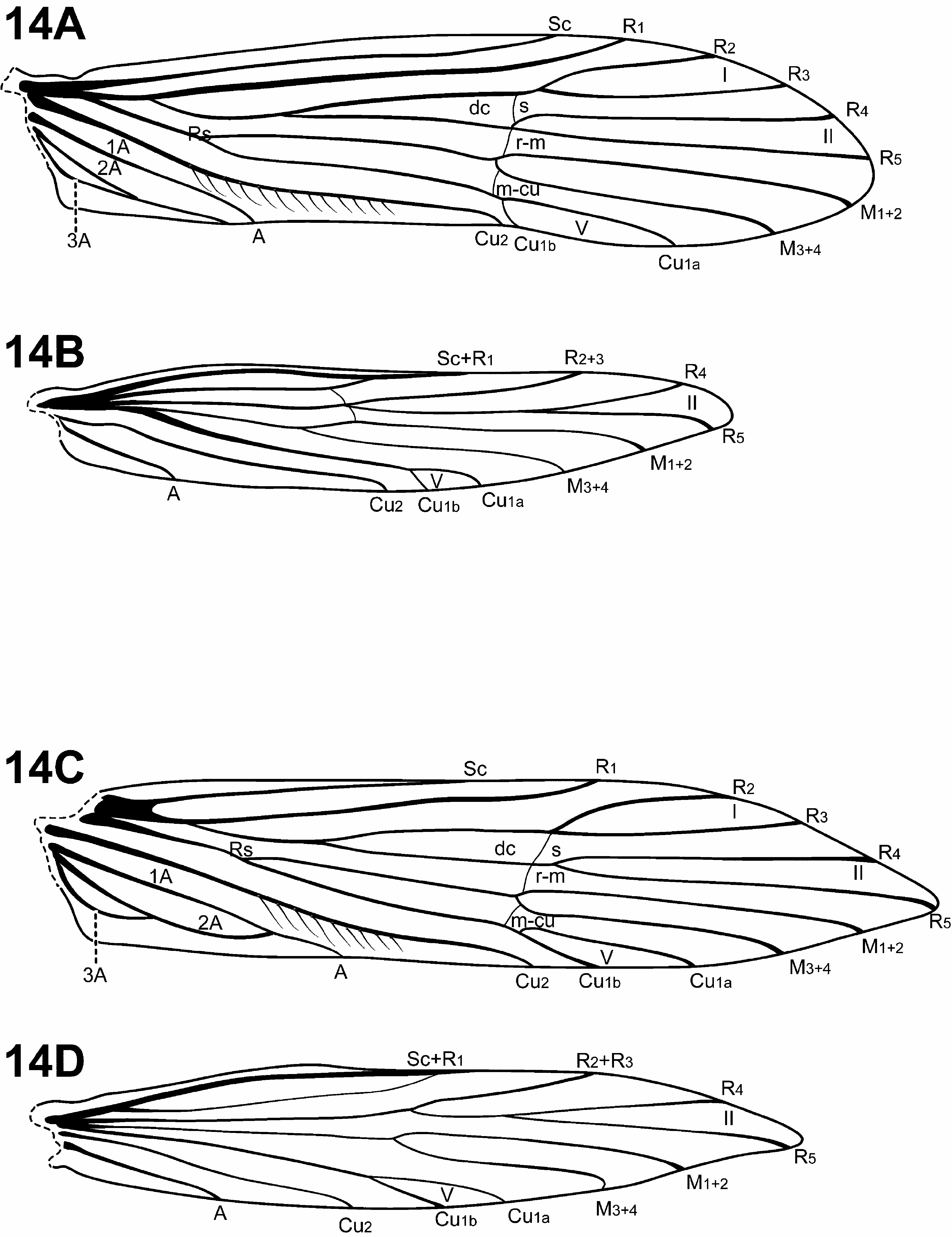
Diagnosis of Padunia ( Figs. 6D View FIGURE 6 , 13 View FIGURE 13 , 14 View FIGURE 14 , 24–27 View FIGURE 24 View FIGURE 25 View FIGURE 26 View FIGURE 27 , 31C View FIGURE 31 ). The male genitalia of Padunia are quite variable among species. Nonetheless, the genus can be diagnosed principally by the structure of the phallobase. The phallobase is very elongate and tubular with a dorsal margin consisting of a sclerotized sheath produced anteriorly. Ventrally, the phallobase is not sclerotized, but highly membranous, or even apparently absent. Another distinctive feature of Padunia is the subtriangular aspect of the anterior margin of segment IX. However, this character is not completely diagnostic for the genus since the segment IX anterior margin of 1 species is slightly rounded ( P. coei ) and another is straight ( P. anakan ). The female genitalia of Padunia are also distinct among protoptilines, with segments VIII, IX, and X fused together dorsally. Another diagnostic feature of Padunia is the presence of a pair of distinct setal patches on the mesoscutellum. These patches differ from the round conspicuous setal warts present in Tolhuaca , but because they may be difficult to see for most species, they may be easily overlooked and have not been rendered or discussed in any any previous species diagnoses or descriptions.
The forewing venation of Padunia is either complete [ P. coei , P. kanikar ( Malicky & Chantaramongkol, 1992) , P. jisunted ( Malicky & Chantaramongkol, 1992) , P. ruangjod ] or incomplete, with apical forks I, II, and V present. While Tolhuaca also has complete forewing venation (plesiomorphic for Trichoptera ), the latter condition (presence of apical forks I, II, and V only), is unique among Protoptilinae . The hind wing of Padunia is most similar to certain species of Itauara , having apical forks II and V present, however the 2 genera can be easily separated based on differences in the male genitalia.
Adult. Body, wings, and appendages nearly uniformly fuscous. Wings often with conspicuous white, broad, transverse marks in anal region and along anastomosis. Head broader than long, vertex rounded, pair of distinct, suboval anterior setal warts, large suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5- segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large subtriangular or suboval pronotal setal warts, occasionally covered with dense scale-like setae. Mesothorax usually wider than long, but longer than wide in P. jeanae , without apparent tegular glands; mesoscutum with pair of elongate anteromesal setal warts, suboval posterolateral warts; mesoscutellum with pair of small, round, distinct setal patches. Forewing ( Figs. 13A, 13C View FIGURE 13 , 14A, 14C View FIGURE 14 ) relatively broad past anastomosis or with margins nearly parallel, apex rounded or subacute, occasionally with scale-like setae. Male occasionally with callosity present in anal and apical costal region of forewing. Forewing venation either complete or incomplete; when incomplete, with apical forks I, II, and V present; Sc and R1 usually distinct along their entire lengths, occasionally intersecting near costal margin; fork I sessile or only slightly petiolate with extremely short stem; fork II sessile or only slightly petiolate with extremely short stem; fork III petiolate, stem about as long as fork; fork IV petiolate, stem about as long as fork; fork V petiolate, or sessile; Cu1 complete, reaching wing margin; Cu1 and Cu2 distinct along their entire lengths; row of erect setae present along Cu2; A3 looped, if present; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein, except in P. falcata and P. phyllis in which the males have highly modified wings with broad half-ellipse-shaped flap along costal margin folding to cover about half wing. Hind wing margins nearly parallel, tapering only slightly past anastomosis; venation variable, either with apical forks II only, or II and III only, or II, III, and V ( P. bilineata species group); Sc and R1 separate or converging near wing margin; A2 absent. Tibial spurs variable, either 0,4,4; 0,4,3; 0,3,3; or 1,4,3; foretibial spur extremely reduced and hair-like. Males occasionally with pair of finger-like lateral processes on sternum V associated with glandular structures ( Fig. 6D View FIGURE 6 ). Sternum VI process present, short and digitate, elongate and digitate, or tooth-like point; apex rounded or attenuate and pointed, usually associated with oblique apodeme posteriorly.
Male genitalia ( Figs. 25–27 View FIGURE 25 View FIGURE 26 View FIGURE 27 ). Terga VII and VIII interstitial region occasionally with glandular structure dorsomesally and dorsolaterally. Segment IX anterior margin usually subtriangular, or rarely, slightly rounded ( P. coei ) or straight ( P. anakan ), posterolateral margin occasionally with lateral processes or lobes; tergum IX well developed, relatively broad; sternum IX usually narrow, occasionally broad ( P. anakan ). Tergum X completely fused to and indistinguishable from segment IX, except in P. anakan in which tergum X appears to be membranous. Inferior appendages present, although often vestigial, or 2 distinct appendages fused to one another basally, attached ventrally to phallus and articulating with segment IX ( P. jeanae ) or fused completely and integrated with segment IX, forming either elongate or short paired ventrolateral processes or single ventromesal process, sometimes bifid. Parameres absent. Phallobase simple, elongate tubular structure, dorsal margin with sclerotized sheath produced anteriorly, with membranous or apparently absent ventral margin. Phallic apparatus occasionally with apicoventral branch and bearing 1 or more apical spines.
Female genitalia ( Fig. 31C View FIGURE 31 ). Truncate posteriorly, not extensible. Abdominal segment VIII syncleritous, anterolateral margin slightly subtriangular or projecting, posterolateral margin often deeply incised. With segments VIII, IX, and X fused, bearing pair of short digitate cerci apically.
Genus Protoptila Banks, 1904
Protoptila Banks, 1904: 215 View in CoL [Type species: Beraea View in CoL ? maculata Hagen, 1861 View in CoL , by original designation].
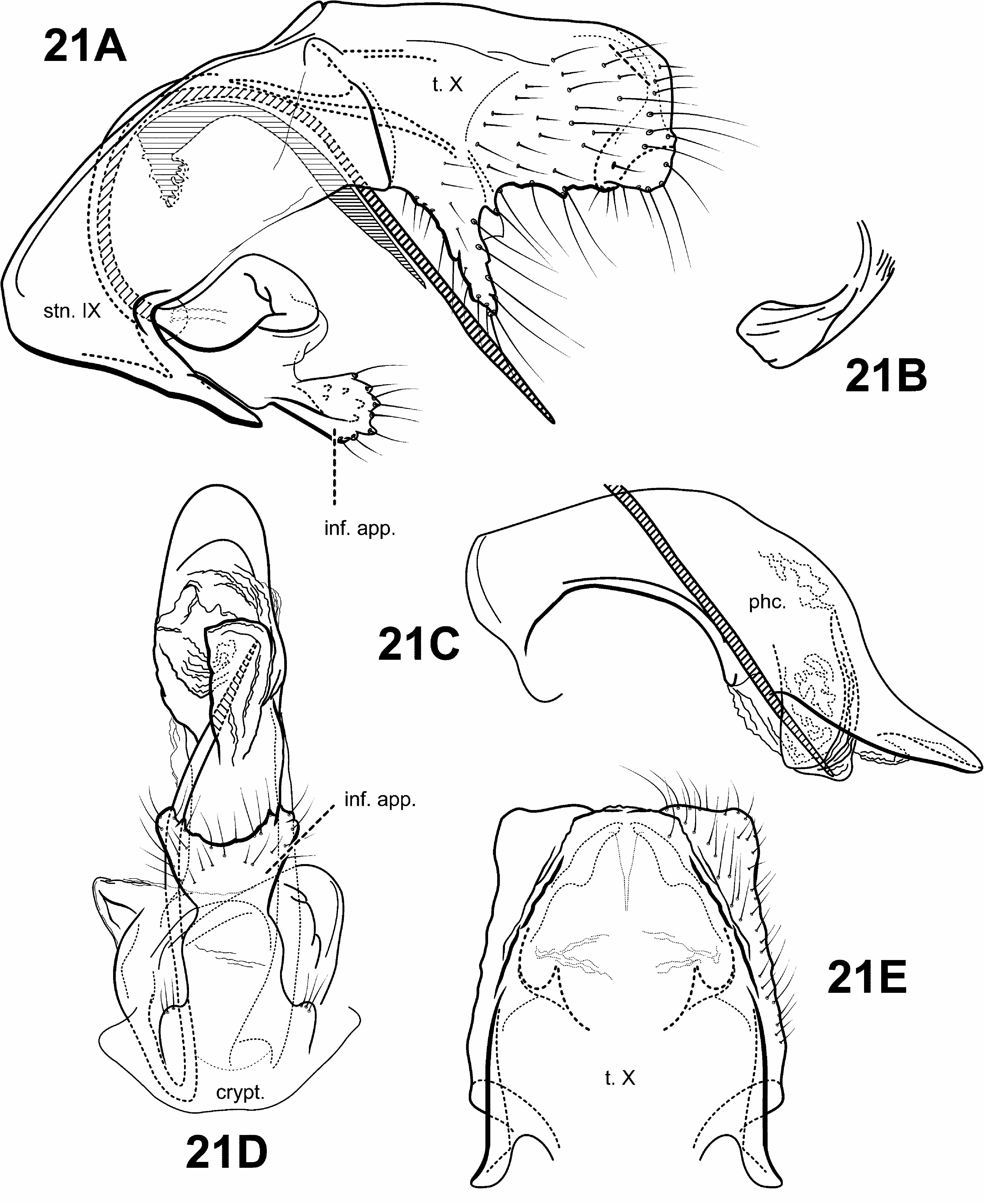
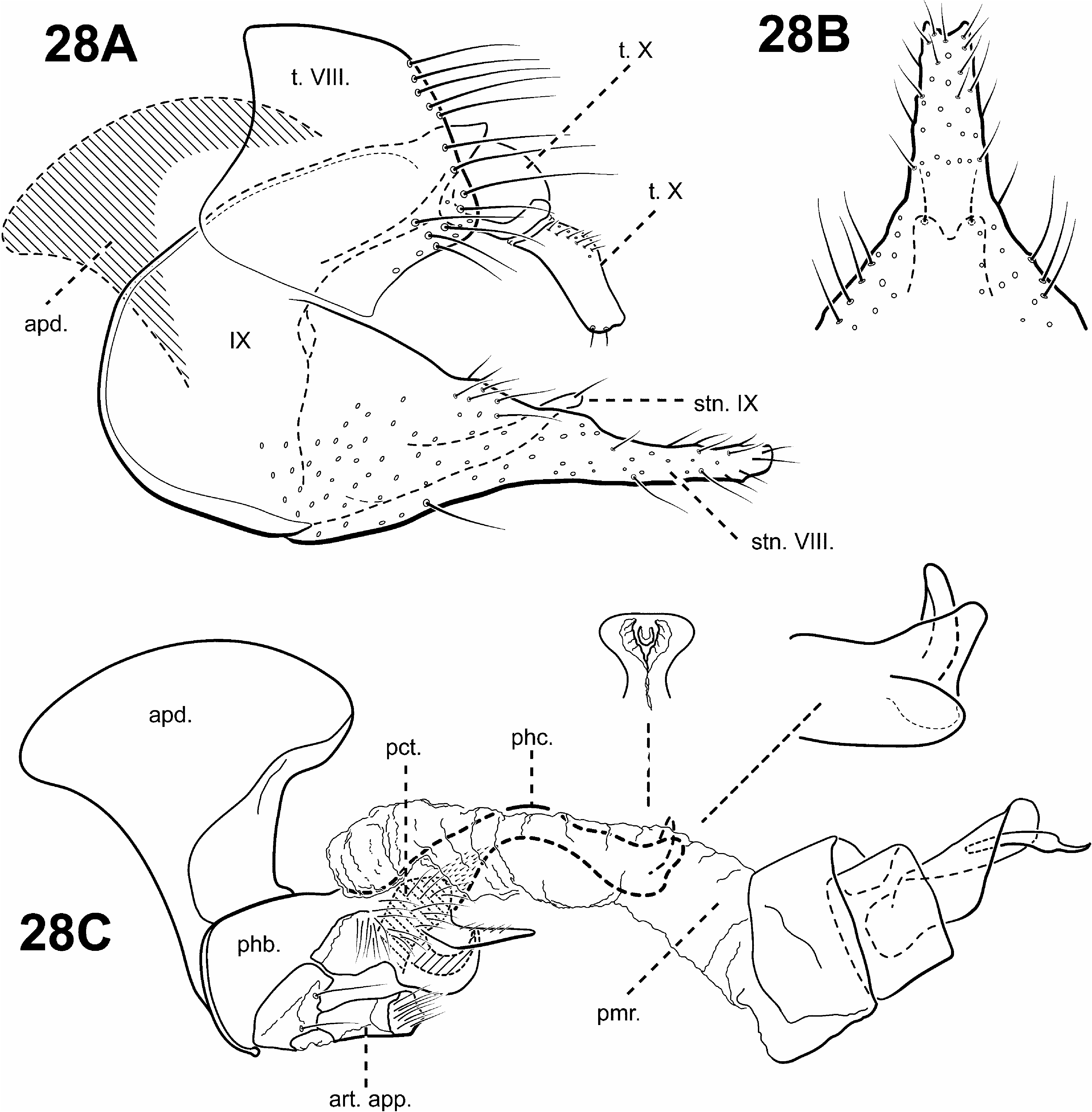
A monophyletic Protoptila View in CoL was strongly supported in all analyses and based on the following synapomorphies suggested by Blahnik and Holzenthal (2006): 1) phallobase with an axe-shaped dorsal apodeme (character 93) ( Fig. 28C View FIGURE 28 ); 2) sternum VIII projecting medially and subtending segment IX to which it is partially fused (character 63) ( Figs. 28A, B View FIGURE 28 ). A 3rd synapomorphy of Protoptila View in CoL includes a ventromesal process on sternum IX projecting posterad (character 72) ( Figs. 28A, B View FIGURE 28 ).
Diagnosis of Protoptila ( Figs. 2C View FIGURE 2 , 3A View FIGURE 3 , 6E View FIGURE 6 , 11C, 11D View FIGURE 11 , 28 View FIGURE 28 ). The genus Protoptila can be recognized by several distinct features of the male genitalia. The dorsum of the phallobase has an enlarged, flattened, often axe-shaped phallic apodeme. Protoptila can also be characterized by the shape of sternum VIII, which projects posterad and subtends sternum IX, to which it is often fused. Additionally, tergum VIII usually has elongate setae along its posterior margin. Tergum X is also distinct among protoptilines, composed of lateral branches, with basal and apical portions. Like Mortoniella , Protoptila also has small, sclerotized, digitate, articulated appendages attached to the phallobase ventrally that fit into modified pockets on the posteroventral portion on the phallobase. However, unlike Mortoniella , Protoptila apparently lacks the associated inferior appendages. The 2 genera can also be separated by differences in the shapes of tergum X and sternum VIII.
The forewing venation is most similar to Mortoniella , with apical forks I, II, and III present, a completely fused Cu1 and Cu2, and 3 anal veins present (although some species of both genera have no 3A). However, the 2 can usually be separated by the row of erect setae, which is positioned far below the Cu2 vein in Protoptila .
Adult. Body, wings, and appendages pale or tawny brown, often intermingled with rufous or golden hairs, tibia and tarsi yellowish brown. Wings often with few pale cream-colored or white hairs, spots, or transverse line along anastomosis and small white specks or spots along apical margin ( Fig. 2C View FIGURE 2 ). Head broader than long, vertex rounded, either 1 distinct pair or 1 divided pair of suboval anterior setal warts, small suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps ( Fig. 6E View FIGURE 6 ) 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly same length as 1st and 2nd segments combined. Prothorax with 2 large, subtriangular or suboval pronotal setal warts, occasionally covered with dense scale-like setae. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval anteromesal setal warts, although occasionally entire region setose with no distinct patch, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 11C View FIGURE 11 ) relatively narrow, with margins nearly parallel, apex oblique. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I, II, and III present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II sessile; fork III petiolate, stem longer than fork; Cu1 complete, reaching wing margin; Cu1 and Cu2 completely fused; row of erect setae present below Cu2; A3 looped, if present; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 11D View FIGURE 11 ) narrow and scalloped past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1,4,4; foretibial spur extremely reduced and hair-like. Sternum VI process present, laterally flattened or subtriangular, apex attenuate and pointed, usually associated with oblique apodeme posteriorly.
Male genitalia ( Fig. 28 View FIGURE 28 ). Preanal appendages absent. Tergum VIII posterior margin with elongate setae. Sternum VIII projecting medially and subtending segment IX. Segment IX anterior margin rounded, posterolateral margin occasionally with lateral processes or lobes; tergum IX usually strap-like or narrow, simple, without processes; mesally, with ventromesal projection directed posterad. Tergum X partially fused to tergum IX, consisting of lateral branches with basal and distal segments, often articulated; dorsomesal margin divided or bifid apicomesally. Inferior appendages absent. Parameres either present or absent; when present, arising laterally from endotheca, often membranous basally, with apical sclerotization and spine, variable in shape. Phallobase not apparently reduced, with enlarged, flattened, often axe-head shaped apodeme dorsally; small pair of articulated digitate, rod-like appendages with membranous apices, associated with modified pockets ventrally; and often with paired, posteriorly projecting processes. Phallic apparatus with varying shape of phallicata, and associated spines and processes. Endophallus membranous, enlarged and convoluted when invaginated, sometimes with sclerotized regions and spines.
Female genitalia. Truncate posteriorly, not extensible. Abdominal segment VIII short, synscleritous, posterolateral margin slightly incised. Sternum VIII often with rounded ventrolateral lobes. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Scotiotrichia Mosely, 1934
Scotiotrichia Mosely, 1934: 160 View in CoL [Type species: Scotiotrichia ocreata Mosely, 1934 View in CoL , by original designation].
Placement of the monotypic genus Scotiotrichia View in CoL remains unresolved. A sister relationship between Scotiotrichia View in CoL and Canoptila View in CoL was weakly supported in the parsimony analysis of the TOTAL MORPH dataset.
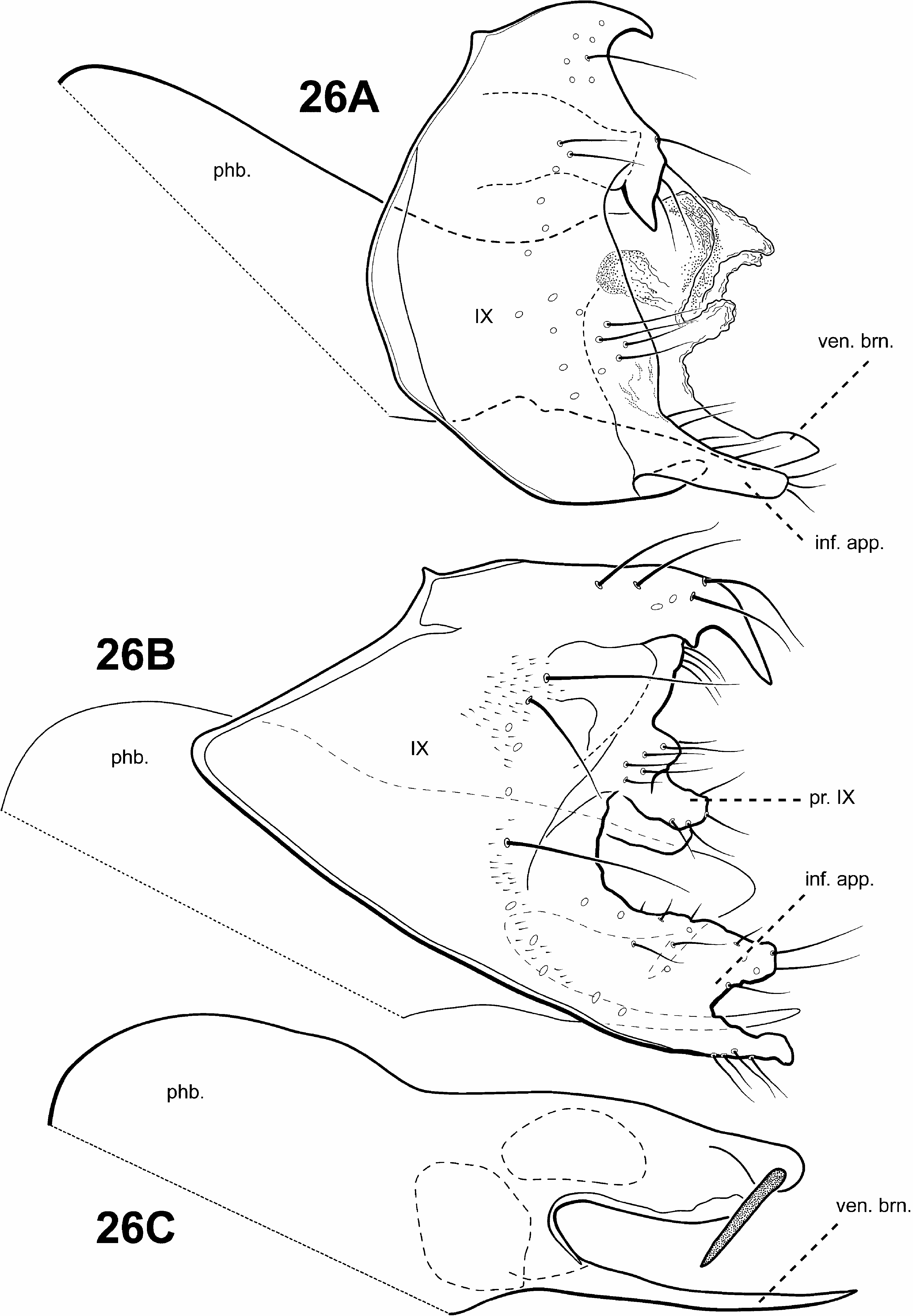
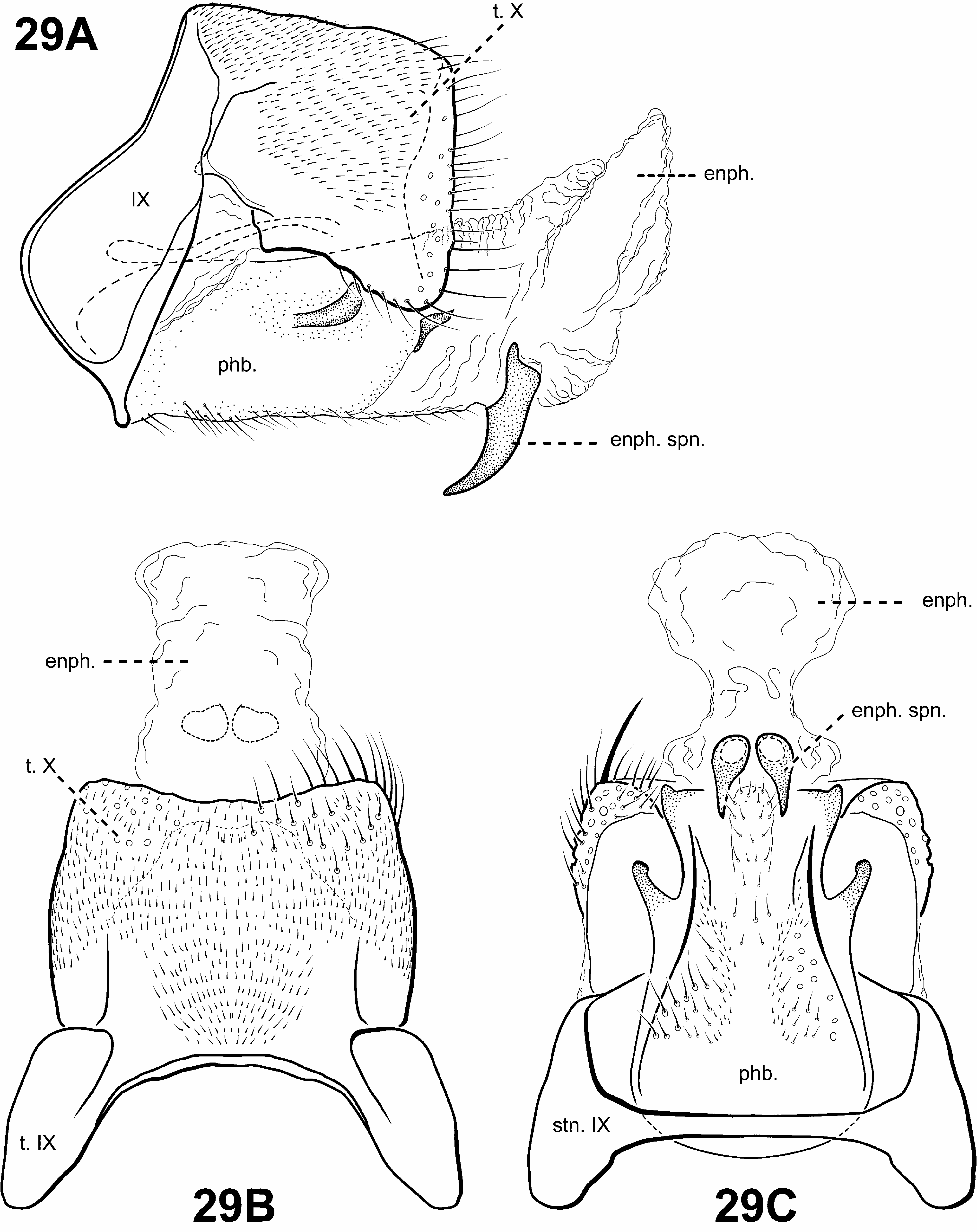
Diagnosis of Scotiotrichia ( Figs. 10C, 10D View FIGURE 10 , 29 View FIGURE 29 ). Scotiotrichia can be recognized by distinct features of the male genitalia and forewing venation. Among protoptilines, the forewing venation of Scotiotrichia is unique, having only apical forks I and II. The male genitalia are simple, with a greatly enlarged and hood-like tergum X. Segment IX is narrow dorsally and straplike ventrally. The phallobase is greatly enlarged, and has a pair of small lateral processes medially, and another pair posterolaterally. The endophallus of Scotiotrichia is large and highly membranous and contains a pair of large, tooth-like processes ventrally. The male genitalia are quite reminiscent of those of Tolhuaca in that both have large phallobases, strap-like sterna IX, and large endophallic membranes. However, the shape of the phallobase differs between the 2 genera: That of Tolhuaca is rounded and much more produced apicomesally whereas in Scotiotrichia , the dorsal and ventral margins are straight. Culoptila and Cariboptila also have enlarged phallobases and strap-like or absent sterna IX, but can easily be differentiated from Scotiotrichia based on their retention of inferior appendages and differences in forewing venation.
Adult. Head broader than long, vertex rounded, pair of distinct, suboval anterior setal warts, large suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5-segmented, 1st and 2nd segments short; 2nd segment bulbous; last 3 segments each nearly as long as 1st and 2nd segments combined. Prothorax with 2 large, subtriangular or suboval pronotal setal warts. Mesothorax wider than long, without apparent tegular glands; mesoscutum with pair of suboval anteromesal setal warts, suboval posterolateral warts; mesoscutellum sparsely setose, without distinct setal warts. Forewing ( Fig. 10C View FIGURE 10 ) relatively narrow, with margins nearly parallel, apex oblique. Male without apparent forewing callosity. Forewing venation incomplete, with apical forks I and II present; Sc and R1 distinct along their entire lengths; fork I sessile; fork II petiolate; Cu1 complete, reaching wing margin; Cu1 and Cu2 intersecting near anastomosis; row of erect setae present below Cu2; A3 absent; crossveins difficult to discern, but apparently forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 10D View FIGURE 10 ) margins nearly parallel, tapering only slightly past anastomosis; apical fork II present; Sc and R1 fused basally; A2 absent. Tibial spurs 1,2,3; foretibial spur extremely reduced and hair-like. Sternum VI process present, short and digitate, apex rounded, associated with strong oblique apodeme posteriorly.
Male genitalia ( Fig. 29 View FIGURE 29 ). Preanal and inferior appendages absent. Segment IX anterior margin rounded, posterolateral margin without lateral process or lobes; tergum IX strap-like, simple, without processes; sternum IX strap-like, mesally, without modification. Tergum X extremely large and hood-like, without processes or lobes, incompletely fused to tergum IX with membrane or lightly sclerotized region ventrolaterally; dorsomesal margin subquadrate. Inferior appendages absent. Parameres absent. Phallobase extremely enlarged, lightly sclerotized with small, stout setae ventrally, medially with small, paired lateral processes, apicomedially with smaller paired processes. Phallic apparatus simple, without spines or processes. Endophallus membranous, enlarged and convoluted when invaginated, bearing single pair of tooth-like downturned spines ventrally.
Female genitalia. Rather elongate posteriorly, but not apparently extensible. Abdominal segment VIII syncleritous, relatively short. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.
Genus Tolhuaca Schmid, 1964
Tolhuaca Schmid, 1964: 336 View in CoL [Type species: Tolhuaca cupulifera Schmid, 1964 View in CoL , by original designation].
Monophyly of Tolhuaca View in CoL was strongly supported in all analyses. Furthermore, a sister relationship to the remaining Protoptilinae View in CoL was found in all analyses, except in the SUBSET COI dataset. Synapomorphies of Tolhuaca View in CoL include a highly inflated, barrel-shaped phallobase and an absence of inferior appendages, although these apomorphies are not unique to the genus.
Diagnosis of Tolhuaca ( Figs. 5A View FIGURE 5 , 6B, 6C View FIGURE 6 , 12A, 12B View FIGURE 12 , 30 View FIGURE 30 , 31A View FIGURE 31 ). The genus Tolhuaca can be recognized by features of the thorax, and male and female genitalia. Among protoptilines, Tolhuaca is distinct in having a pair of conspicuous round setal warts on the mesoscutellum, the plesiomorphic condition in Trichoptera . Padunia also has setal warts on the mesoscutellum, but they differ from those of Tolhuaca in being more patch-like and less conspicuous. The phallobase of Tolhuaca is extremely enlarged and barrel-shaped, with a sclerotized projection apicomesally. An additional distinct feature includes the large, tubular endophallus, which is highly membranous and contains several sclerotized spines and other internal structures. The male genitalia are similar to those of Scotiotrichia , Cariboptila , and Culoptila —each have enlarged phallobases and reduced or absent sterna IX. However, these genera can be separated based on differences in the shape of the phallobase and wing venation. The female genitalia are also unique among Protoptilinae . They are elongate and oviscapt, and have 2 pairs of rod-like internal apodemes. The female genitalia of Scotiotrichia are also rather elongate, but do not appear to be oviscapt, and lack the rod-like apodemes. The forewing venation of Tolhuaca is complete, like that of some species of Padunia , yet these genera are easily separated by differences in the shape of sternum IX in the male genitalia.
Adult. Tibia and tarsi yellowish brown. Head ( Fig. 5A View FIGURE 5 ) broader than long, vertex rounded, with pair of small anteromesal setal warts, pair of distinct, elongate anterior setal warts, large suboval posterior setal warts, suboval or triangular and bulging posterolateral setal warts. Ocelli present. Antennal scape less than or equal to 2 times length of pedicel. Maxillary palps 5-segmented, 1st and 2nd segments short with elongate setae apically; 2nd segment bulbous; last 3 segments each nearly as long as 1st and 2nd segments combined. Prothorax ( Fig. 5A View FIGURE 5 ) with 2 large subtriangular or suboval pronotal setal warts. Mesothorax ( Fig. 5A View FIGURE 5 ) longer than wide, without apparent tegular glands; mesoscutum with pair of elongate anteromesal setal warts, suboval posterolateral warts; mesoscutellum with pair of small, round, distinct setal warts. Forewing ( Fig. 12A View FIGURE 12 ) relatively broad past anastomosis, apex rounded, with erect or retrorse setae along some veins, most noticeably along Cu2. Male without apparent forewing callosity. Forewing venation complete; Sc and R1 distinct along their entire lengths; fork I petiolate, but with extremely short stem, or sessile; fork II sessile; fork III petiolate, stem shorter than fork; fork IV petiolate, stem shorter than fork; fork V sessile; Cu1 complete, reaching wing margin; Cu1 and Cu2 distinct along their entire lengths; row of erect setae present along Cu2; A3 looped; crossveins forming relatively linear transverse cord; discoidal cell longer than Rs vein. Hind wing ( Fig. 12B View FIGURE 12 ) broad past anastamosis; apical forks II, III, and V present; Sc and R1 fused basally or converging near wing margin; A2 present, looped. Tibial spurs 1,4,4; foretibial spur extremely reduced and hair-like ( Fig. 6B, 6C View FIGURE 6 ). Sternum VI process present, thumb-like and prominent or elongate and digitate, apex rounded, associated with strong oblique apodeme posteriorly.
Male genitalia ( Fig. 30 View FIGURE 30 ). Preanal and inferior appendages absent. Segment IX anterior margin rounded; tergum IX well developed, relatively broad, simple, without processes; sternum IX strap-like. Tergum X completely fused to tergum IX but with membranous connection visible; dorsomesal margin divided or bifid apicomesally; dorsolateral margin without processes; ventrolateral margin with small, irregular, paired setose lobes. Parameres absent. Phallobase extremely enlarged, barrel-shaped, lightly sclerotized with small, stout setae, without processes, produced and sclerotized apicomesally. Phallic apparatus simple, without spines or processes. Endophallus membranous, greatly enlarged, and rather tubular when evaginated, with sclerous spines or rod-like structures internally.
Female genitalia ( Fig. 31A View FIGURE 31 ). Extensible oviscapt. Abdominal segment VIII syncleritous, about as wide as long. Internally, with 2 pairs of long, slender, sclerotized, rod-like apodemes arising from lateral margins of segments VIII and IX and extending cephalad to segments VI and VII, respectively. Segments IX and X closely associated, with pair of small digitate cerci dorsolaterally.


















No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Protoptilinae Ross, 1956: 149
| ROBERTSON, DESIREE R. & HOLZENTHAL, RALPH W. 2013 |
Temburongpsyche
| Malicky, H. 1995: 15 |
Cubanoptila
| Botosaneanu, L. & Sykora, J. 1973: 383 |
Cariboptila
| Flint, O. S. Jr. 1964: 17 |
Campsiophora
| Flint, O. S. Jr. 1964: 14 |
Nepaloptila
| Kimmins, D. E. 1964: 37 |
Tolhuaca
| Schmid, F. 1964: 336 |
Merionoptila
| Schmid, F. 1959: 482 |
Protoptilinae
| Ross, H. H. 1956: 149 |
Matrioptila
| Ross, H. H. 1956: 164 |
Culoptila
| Mosely, M. E. 1954: 336 |
Uenotrichia
| Tsuda, M. 1942: 228 |
Canoptila
| Mosely, M. E. 1939: 218 |
Mexitrichia
| Mosely, M. E. 1937: 158 |
Scotiotrichia
| Mosely, M. E. 1934: 160 |
Padunia
| Martynov, A. B. 1910: 425 |
Mortoniella
| Ulmer, G. 1906: 95 |
Protoptila
| Banks, N. 1904: 215 |
Itauara Müller, 1888: 275
| Mosely, M. E. 1939: 219 |
| Muller, F. 1888: 275 |