
Persicaria hydropiper, L.
|
publication ID |
https://doi.org/ 10.1016/j.phytochem.2014.10.001 |
|
DOI |
https://doi.org/10.5281/zenodo.10570181 |
|
persistent identifier |
https://treatment.plazi.org/id/1A2A87A6-FF9A-FFA7-4C70-BDD0B21964C3 |
|
treatment provided by |
Felipe |
|
scientific name |
Persicaria hydropiper |
| status |
|
2.2. Accumulation pattern of secondary metabolites during flower and leaf development in P. hydropiper View in CoL
To investigate the relative abundance of the secondary metabolites possibly involved in defence throughout development, three stages of flowers and four of leaves were analysed for their chemical composition. Table 4 View Table 4 shows how all drimane sesquiterpenoids were most abundant in Flower stage B. The leaf content of these chemicals was 13% or less, compared to flowers, and the trend was that smaller leaves contained more drimanes. This is possibly due to a relative decrease in cavity density per surface area as leaves expand, as is the case with trichomes of some plant species ( Ascensao and Pais, 1987). By MANOVA differences between flowers and leaves were significant for all compounds, but generally not between stages. However, a paired t -test across all compounds per stage did reveal some significant differences between stages ( Table 4 View Table 4 ).
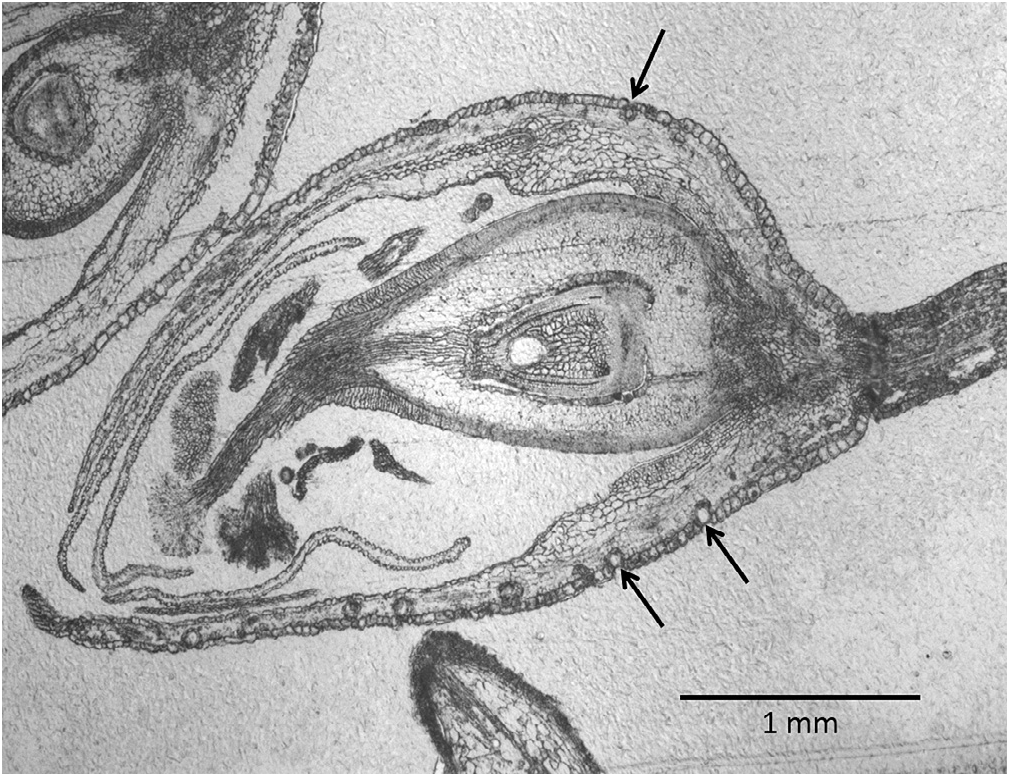
The accumulation patterns of different compounds throughout flower development were grouped together by means of a similarity analysis in order to point to common biosynthetic pathways, common storage tissues like cavities or common biological roles or physical properties. The similarity matrix in Fig. 2 View Fig shows how flower derived compounds of P. hydropiper cluster. The phylogenetic tree, obtained with the Unweighted Pair Group Method with Arithmetic mean (UPGMA) hierarchical clustering method, shows four distinct groups when half of the clustering distance is considered as a relevant cut-off point. The two minor groups seen at the left hand side and bottom of the matrix represent non-terpenoids (others) and diterpene neophytadiene isomers, with the exception of drimenol. Drimenol is an intermediate in the biosynthesis of polygodial and would not be expected to be stored in the cavities, which are not biosynthetically active, but rather in surrounding cells, which may produce the contents of the cavities. Therefore, an accumulation pattern of drimenol different from polygodial is expected. The three major drimane sesquiterpenes cluster together and belong to the largest group, which, next to bornyl acetate and dihydro-a- ionone, contains 12 sesquiterpenes. This could indicate that all these compounds are stored in the cavities ( Fig. 1 View Fig ). The other large cluster contains the monoterpene limonene with five other sesquiterpenes. They are characterised by lower levels in later flower stages ( Table 4 View Table 4 ). Possibly these are regulated differently, more volatile and/or not localised in the valvate glands.
2.3. Volatile emissions from P. hydropiper View in CoL
In our investigation, we also analysed the headspace of P. hydropiper . We focussed only on this species, as it had a far higher content of secondary metabolites in the extracts. The volatile blend emitted by leaves and flowers of P. hydropiper contained eight major compounds which were, identified like the compounds in the extracts, by matching the library hits with the known and observed retention indices ( Table 5 View Table 5 ). The six sesquiterpenes in the headspace more or less reflect the relative contents in the extracts of flowers and leaves. No drimanes were detected in the headspace, presumably because their predicted boiling point (~ 323 °C for polygodial) is at least 50° higher compared to that of other sesquiterpenes (e.g. 261 °C for zingiberene), which results in a 228-fold lower vapour pressure and thus volatility (0.000107 vs. 0.0245 mm Hg at 25 °C for polygodial and zingiberene, respectively) (Values were generated using the US Environmental Protection Agency’s EPISuite™; information retrieved at www.chemspider.com). The monoterpene limonene was much more dominant in the headspace of flowers than in the extract, presumably due to its high volatility.
3. Conclusions
In this study, we compared the chemical profiles of three species of smartweed. In all three species, the flowers contained higher amounts and more types of compounds compared with leaves of the same plant. The vast majority of the compounds identified were sesquiterpenes, five of which were drimanes. We observed that P. hydropiper produced the highest amounts of sesquiterpene secondary metabolites, while its congeners, by comparison, accumulated at least 100–500 times less in flowers, and 15– 100 times less in leaves. This differential investment in making secondary metabolites might reflect differences in defence strategies or needs between the three species of smartweed. Interesting, in that respect, is the observation that in P. maculosa , in both leaves and flowers, there seems to be selective loss (or lack of selected gain) of drimanes and most sesquiterpenes except for (E)-b- farnesene and germacrene A. According to the Optimal Defence Theory (ODT), the chemical-defence needs of any part of the plant are determined by value and vulnerability ( McCall and Fordyce, 2010). In this frame, young developing tissues are very vulnerable to predation by herbivores. This could explain the higher quantity of polygodial found in young leaves as well as in the tepals of those flower heads with a young developing fruit, although it does not explain why three closely related species, occurring in the same habitats have diverged so much with respect to drimane-based defences. Quite possibly the vulnerability of the other two species to whatever attacks P. hydropiper is less, so that there has been not sufficient value to drimane accumulation to balance the cost associated with producing it. A pattern similar to that of P. hydropiper accumulation of polygodial (much higher in flowerheads than in leaves) has been observed in the unrelated shrub Pseudowintera colorata (Raoul) , where berries contained roughly 100 times more polygodial and 9-deoxymuzigadial compared to leaves ( Larsen et al., 2007). Those authors speculate, that the presence of those two pungent sesquiterpene dialdehydes might exert a protective function against non-specialist herbivores. Considering the broad action of polygodial against mammals, insect pests and microbial pathogens, we would argue that ecological studies with genotypes that are differential in the accumulation of drimanes can provide the necessary evidence for this.

| A |
Harvard University - Arnold Arboretum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |