
Esophyllas, Prentice, Thomas R. & Redak, Richard A., 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.213135 |
|
DOI |
https://doi.org/10.5281/zenodo.6180420 |
|
persistent identifier |
https://treatment.plazi.org/id/1B4F87C7-FFAA-FFEC-FF5B-46623EE1FAA6 |
|
treatment provided by |
Plazi |
|
scientific name |
Esophyllas |
| status |
gen. nov. |
Esophyllas View in CoL new genus
Type species. Esophyllas vetteri new species
Etymology. The name is derived from the Greek language and is feminine in gender, the prefix eso- meaning ‘within’ and phyllas meaning ‘bed of leaves’, referring to the habitat from which almost all specimens were collected, more specifically, oak leaf litter.
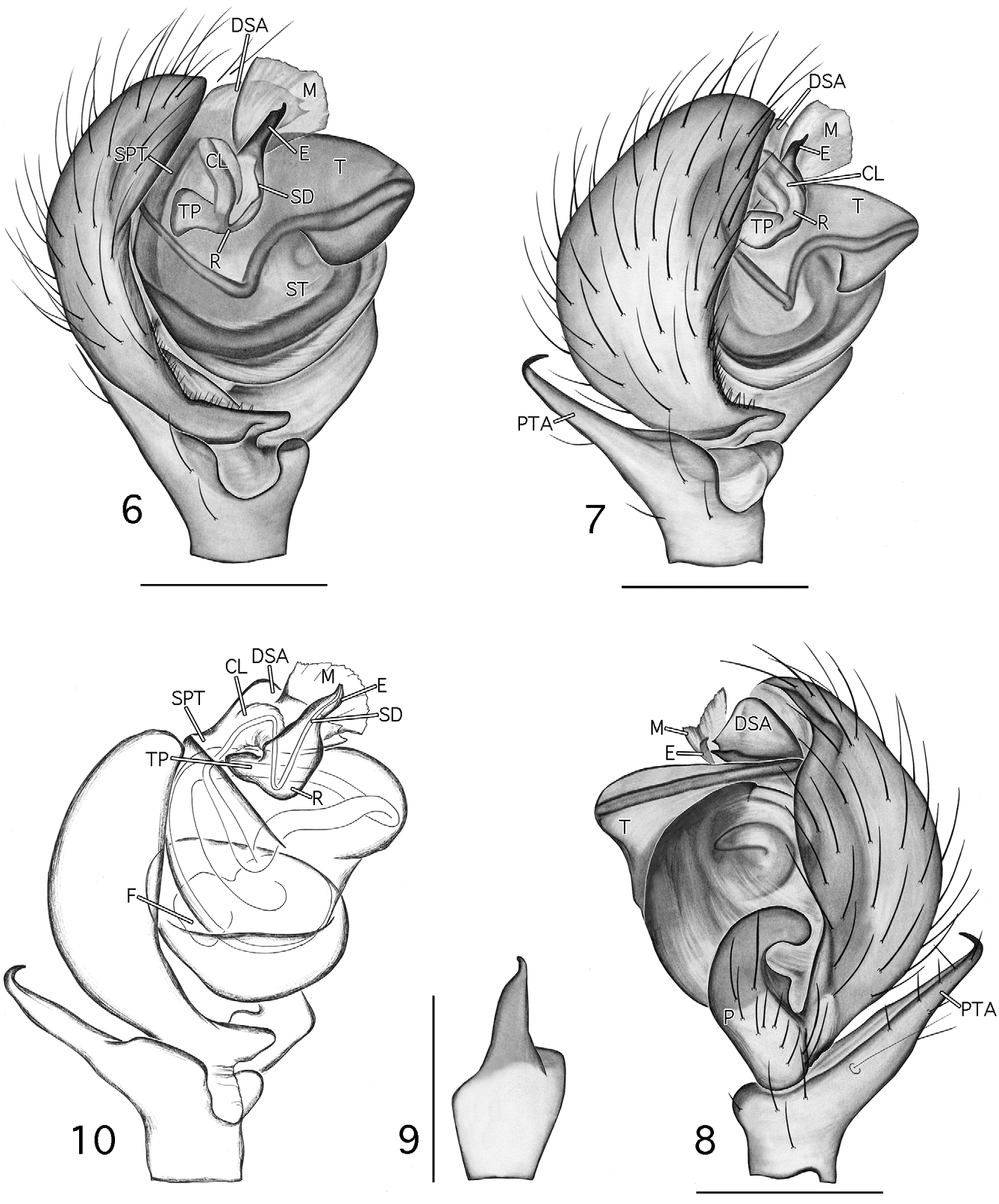
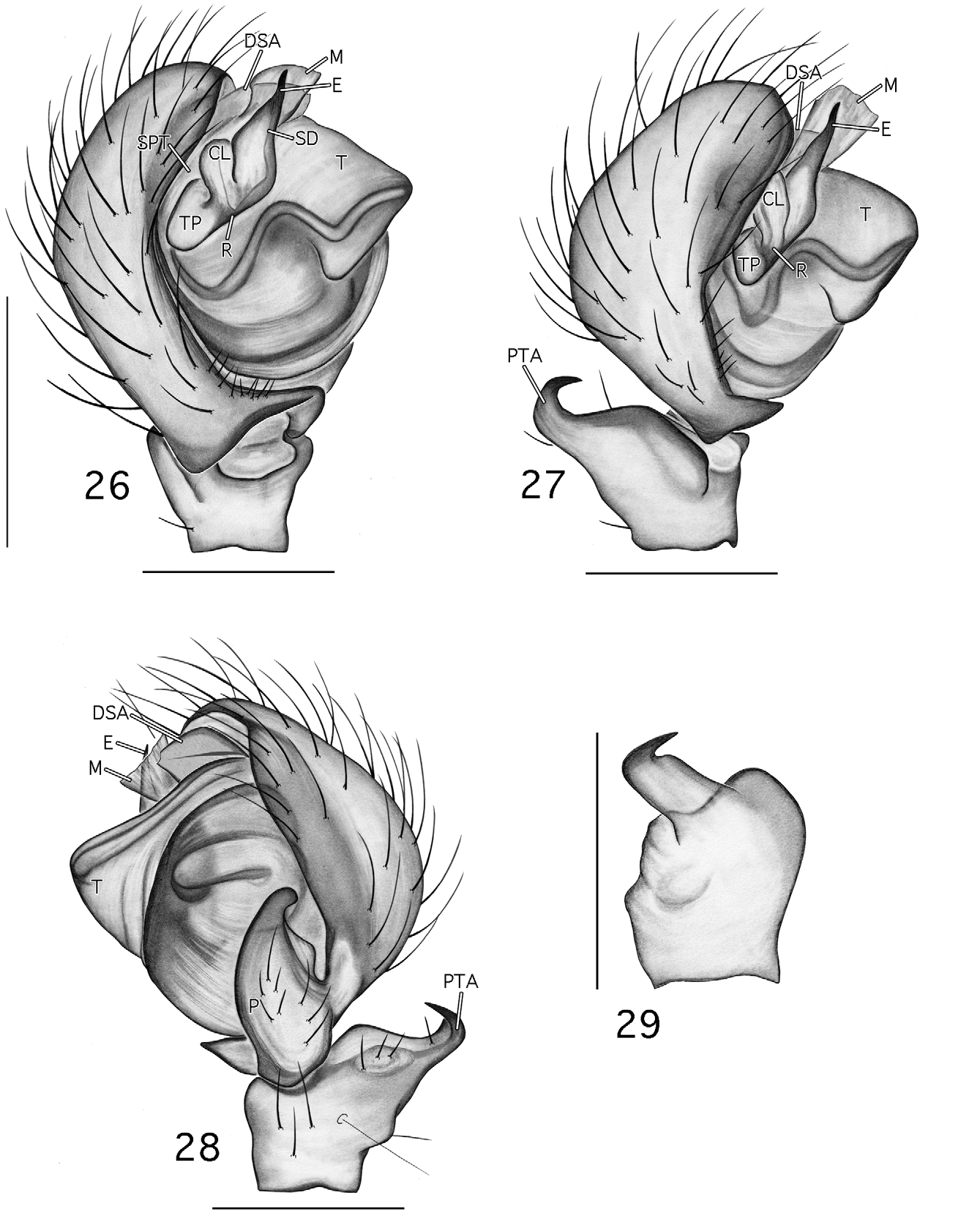
Diagnosis. Esophyllas n. gen. is composed of two small desmitracheate (taenidia absent; Blest 1976) species with short emboli and is distinguished from all genera in the ‘ Tapinocyba group’ ( Millidge 1977; following current placement: Tapinocyba , Phlattothrata , Crosbyarachne Charitonov 1937 , Ceratinops Banks , Mecynargus Kulczin’ski, Semljicola Strand , Thyreosthenius Simon , Acartauchenius Simon , Trichopternoides thorelli (Westring) , and Erigonoplus Simon ) and Scirites by the following male characters: presence of post-PME cephalic lobe (also in Phlattothrata and Tapinocyba prima Dupérré & Paquin 2005 ), absence of the protegulum, a strong proximal to distal contraction of the tegulum ( Figs 8 View FIGURES 6 – 10 , 28 View FIGURES 26 – 29 : T), a tubular sclerotized column ( Figs 6 View FIGURES 6 – 10 , 26 View FIGURES 26 – 29 : CL; also present in P. parva ), sperm duct looped over the embolic division (ED) prior to entering the radix on the proximomesal side of the ED ( Figs 7 View FIGURES 6 – 10 , 27 View FIGURES 26 – 29 : R), entry further posteriad and closer to the middle of the bulb ( Figs 6 View FIGURES 6 – 10 , 26 View FIGURES 26 – 29 ; loop and entry similar in Crosbyarachne silvestris ( Georgescu 1973) but column is membranous and carries an embolic membrane), tibial spine arrangement of 2-2-1-1 (vs. 1-1-1-1, 2-2-2-1, or in Erigonoplus , 2-2-1-1), proximal to distal orientation of the paracymbium (length greater than width, Figs 8 View FIGURES 6 – 10 , 28 View FIGURES 26 – 29 : P; similar in P. parva ), and presence of a flat triangular process projecting from the dorsal edge of the paracymbium ( Figs 8 View FIGURES 6 – 10 , 28 View FIGURES 26 – 29 : P) and a heavily sclerotized refexed barb forming the terminus of the tibial apophysis ( Figs 7, 8 View FIGURES 6 – 10 , 27, 28 View FIGURES 26 – 29 : PTA). The latter two characters are considered to be synapomorphies of Esophyllas n. gen.
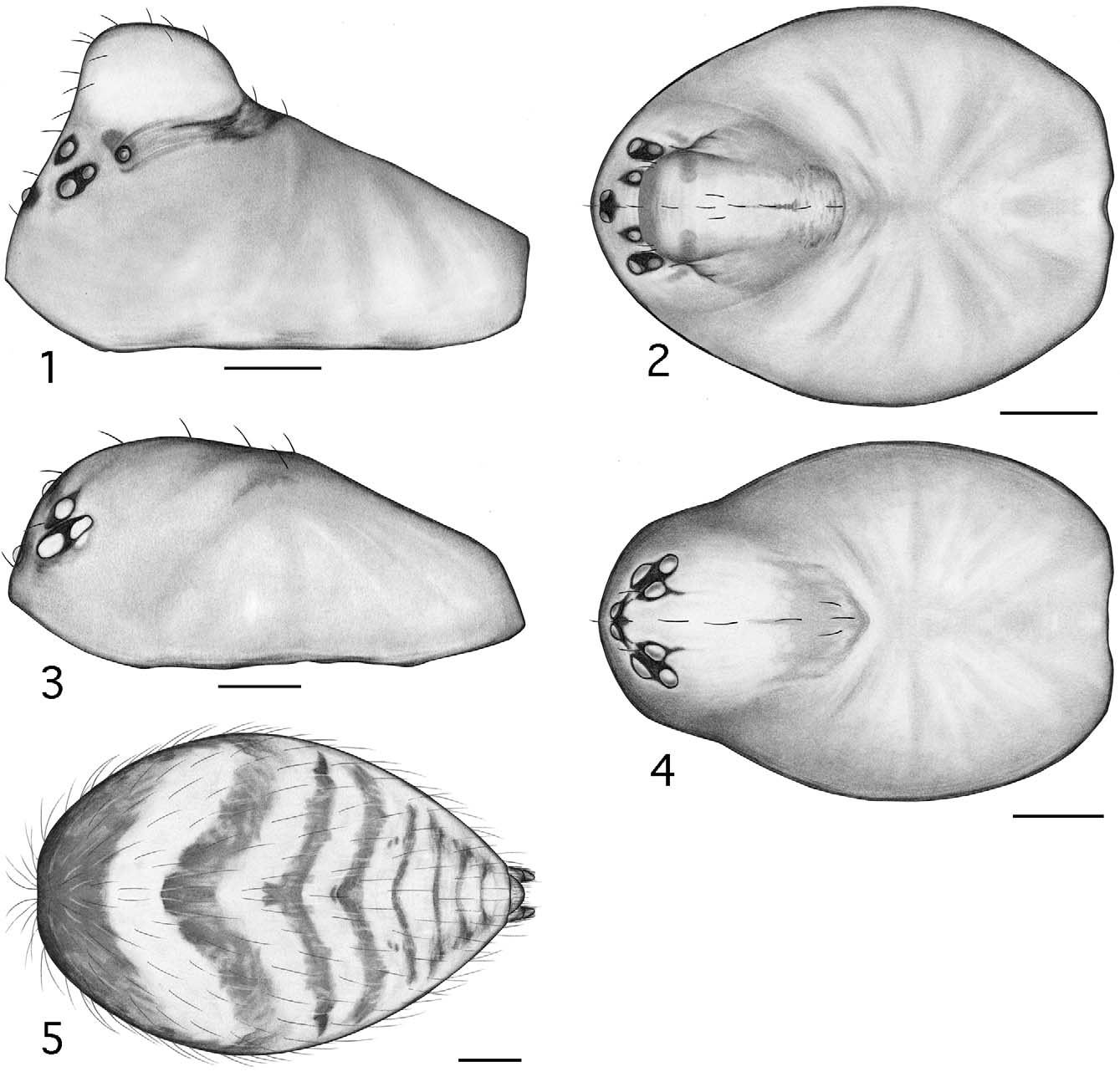
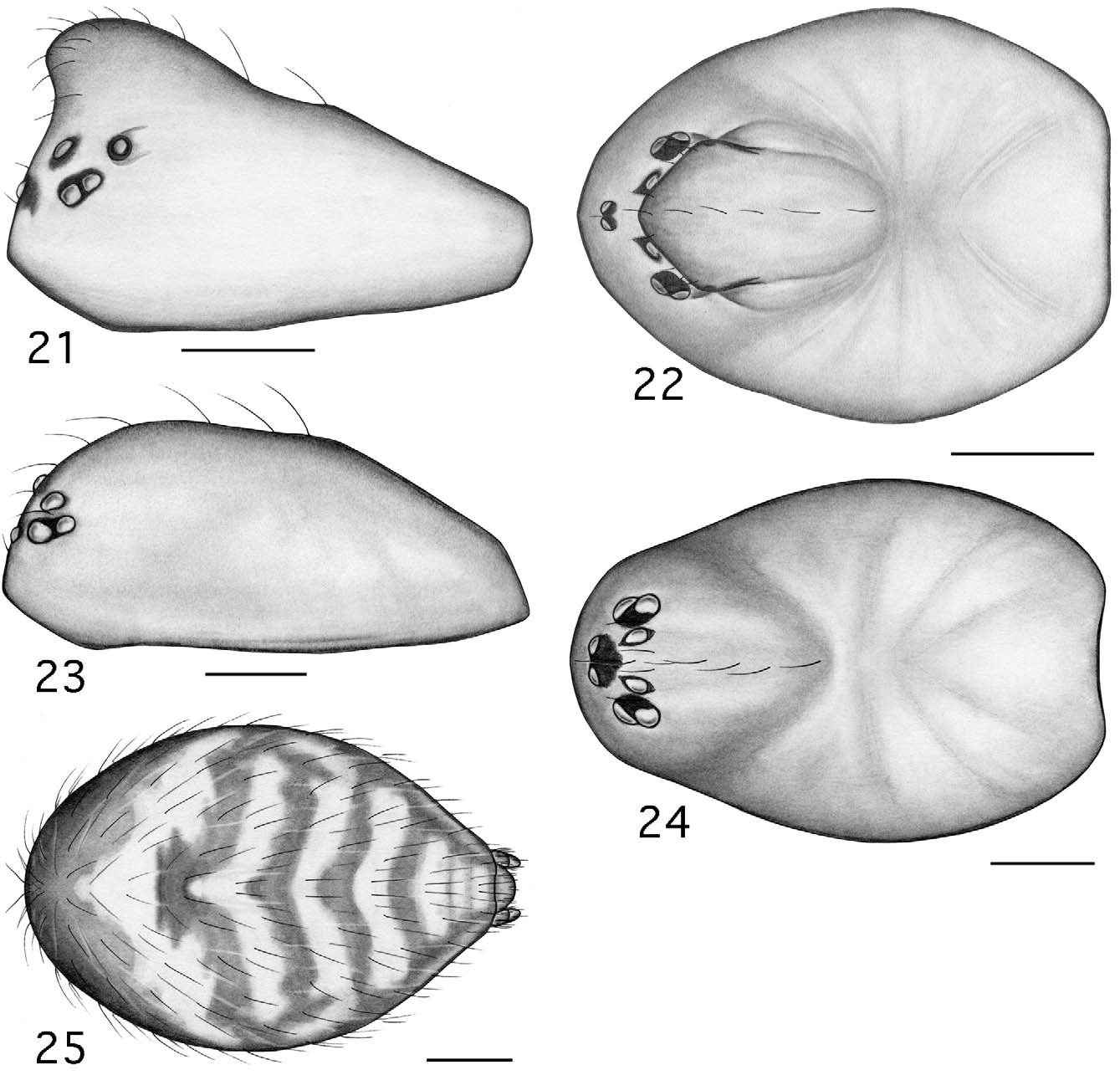
Males are further separated from those of four of the above genera ( Tapinocyba , Phlattothrata , Crosbyarachne , Scirites ), with which Esophyllas n. gen. is believed to be more closely related, by the following: from Tapinocyba by the absence of pectinate tarsal claws, a dorsal seta on the palpal patella, a prolateral trichobothria on the palpal tibia, and a sudden constriction in the diameter of the sperm duct; from P. p ar v a by the absence of an anterior radical process, an embolic membrane (SEM imaging necessary to confirm attachment points in P. pa r v a; not a permitted option for borrowed material), pectinate tarsal claws, and a prolateral trichobothria on the palpal tibia ( Figs 7 View FIGURES 6 – 10 , 27 View FIGURES 26 – 29 ), and by the position of the lateral sulci and prosomal pits ( Figs 1 View FIGURES 1 – 5 , 21 View FIGURES 21 – 25 ; compare to Paquin & Dupérré 2003; fig 1273, P. flagellata (Emerton)) ; from Crosbyarachne by the presence of a short straight embolus (vs. long and coiled), absence of an embolic membrane, a sudden constriction in the diameter of the sperm duct, pectinate tarsal claws, and a prolateral trichobothrium on the palpal tibia, and by the patterned abdomen; from Scirites by the absence of a row of strong megaspines on metatarsus I and papillae on the tegulum, pleurites and sternite of the male pedicel not juxtaposed, the presence of lateral sulci, prosomal pits, and patterned abdomen ( Figs 1, 5 View FIGURES 1 – 5 , 21, 25 View FIGURES 21 – 25 ).
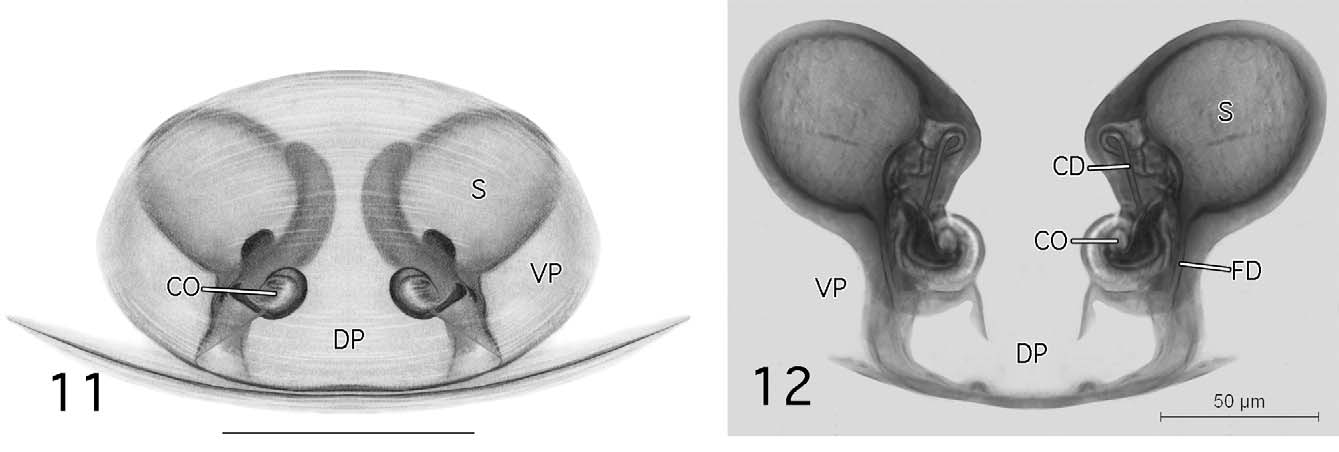
Females are most easily confused with Spirembolus pusillus Millidge and S. novellus Millidge (refer to Millidge 1980b) that have similar epigynal morphology, abdominal patterning and distributions (either overlapping or closely adjacent) and can be distinguished by the smaller size, greater width between the ectal margins of the spermathecae than the width of the dorsal plate ( Figs 11 View FIGURES 11 – 12 , 30 View FIGURES 30 – 31 : S, DP), convolution of the copulatory ducts only on the mesal side of the spermathecae ( Figs 12 View FIGURES 11 – 12 , 31 View FIGURES 30 – 31 : CD), copulatory ducts with only a single loop prior to entering the spermathcae on the mesal side ( Figs 12 View FIGURES 11 – 12 , 31 View FIGURES 30 – 31 : CD), TmI 0.30–0.40, and tibial spine arrangement of 2-2-1-1.
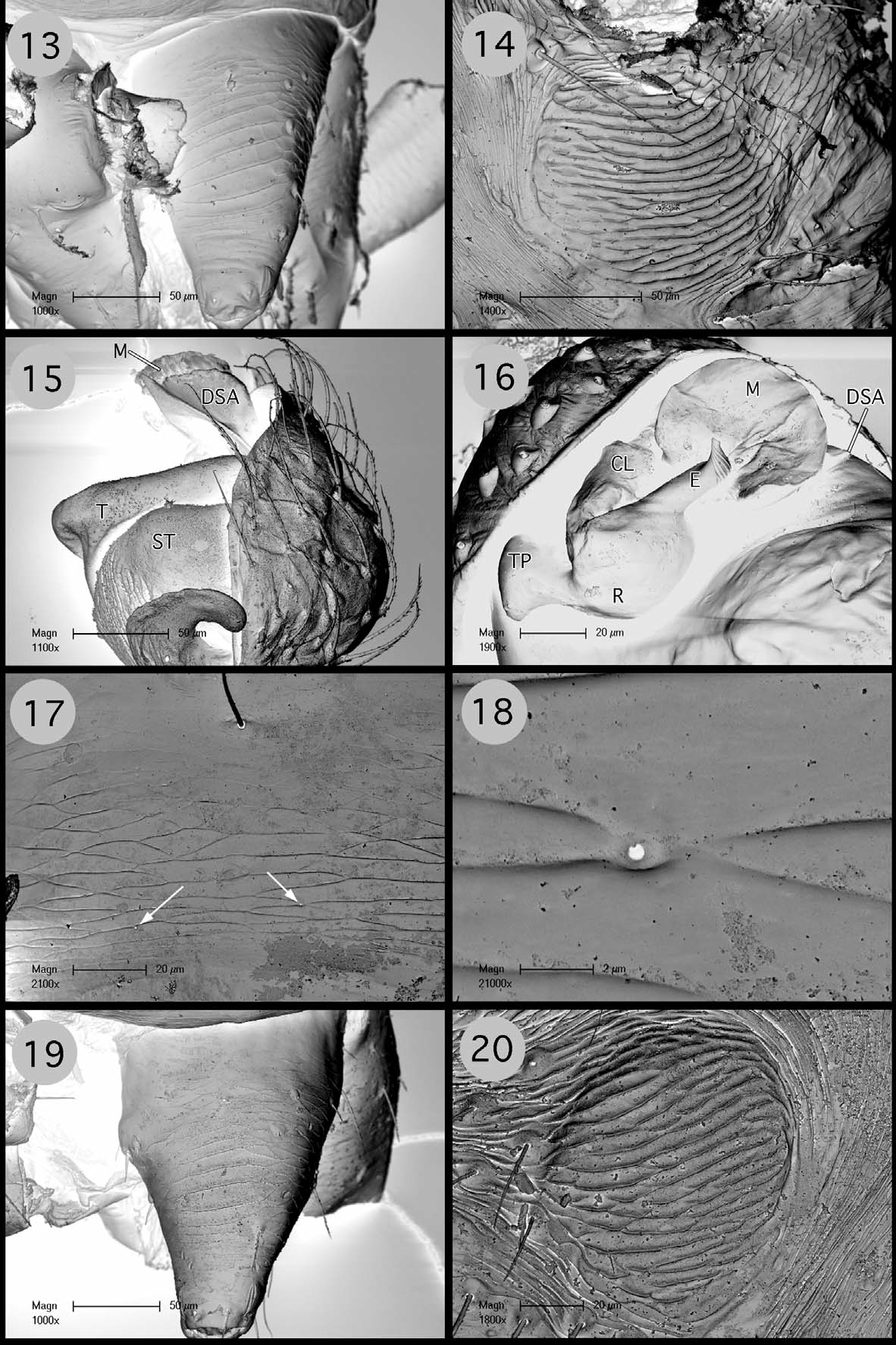
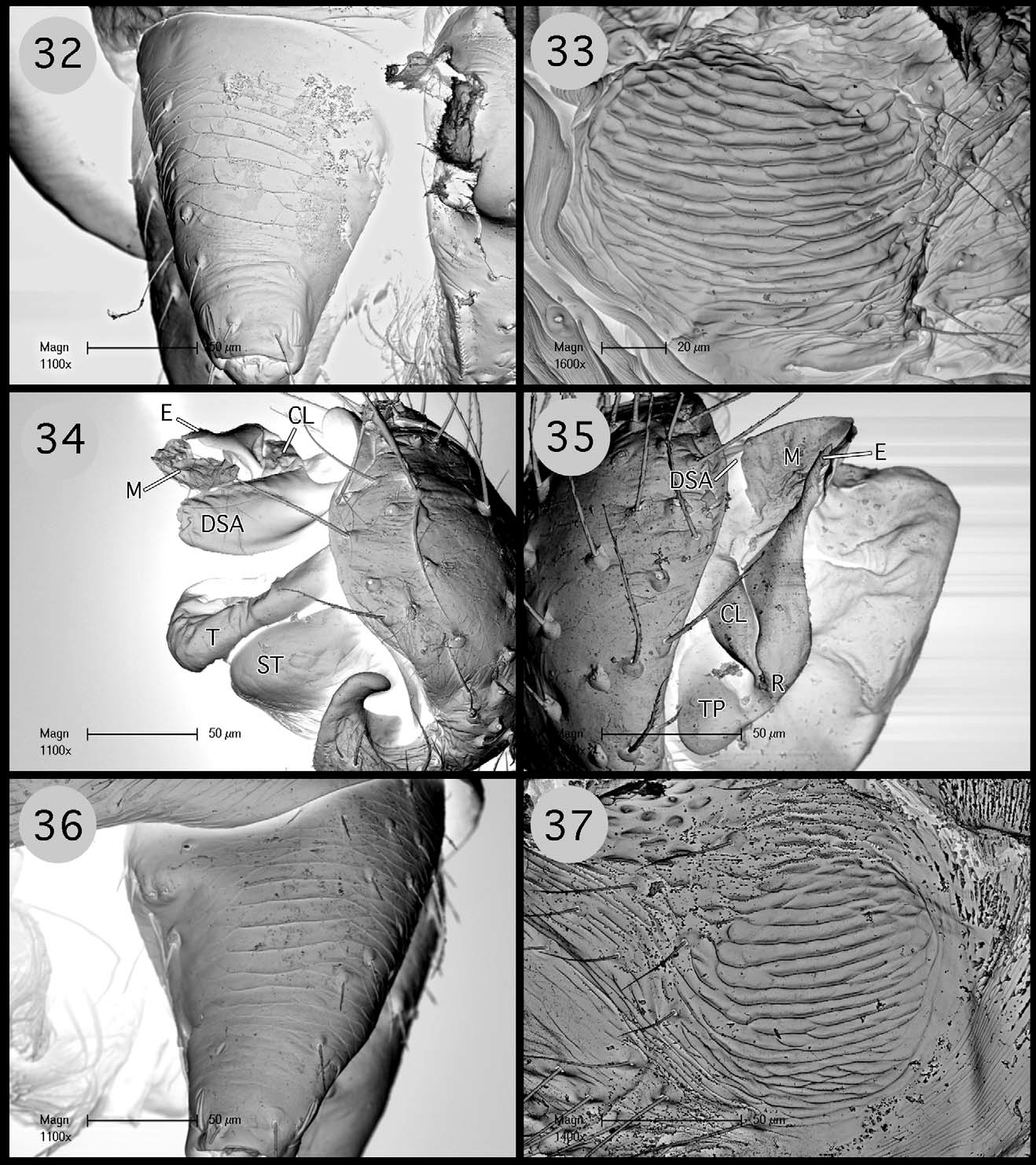
Description. Small desmitracheate (taenidia absent; Blest 1976) erigonines with males ranging in total length from 1.05–1.25 and females from 1.15–1.60, TmI 0.31–0.41, TmIV absent, tibial spines 2-2-1-1, stridulatory mechanisms on both the chelicerae ( Figs 13, 19 View FIGURES 13 – 20 , 32, 36 View FIGURES 32 – 37 ) and booklung plates ( Figs 14, 20 View FIGURES 13 – 20 , 33, 37 View FIGURES 32 – 37 ), yellowishorange color of the carapace, chelicerae, sternum, and legs, and a dark abdominal herringbone pattern ( Figs 5 View FIGURES 1 – 5 , 25 View FIGURES 21 – 25 ). Males are further recognized by the post-PME cephalic lobe with lateral sulci and prosomal pits ( Figs 1 View FIGURES 1 – 5 , 21 View FIGURES 21 – 25 ), the ED consisting of a short straight embolus with spiraled and grooved terminus ( Figs 16 View FIGURES 13 – 20 , 35 View FIGURES 32 – 37 : E), very short tailpiece ( Figs 6 View FIGURES 6 – 10 , 26 View FIGURES 26 – 29 : TP), with the sperm duct looped over the ED through a sclerotized tubular column prior to entering the radix on the mesal surface of the ED near the middle of the bulb ( Figs 7 View FIGURES 6 – 10 , 27 View FIGURES 26 – 29 : CL, R). The tegulum strongly is compressed between proximal and distal margins ( Figs 8 View FIGURES 6 – 10 , 28 View FIGURES 26 – 29 : T) and both the protegulum and embolic membrane are absent; the membrane that is present is articulated with only the distal suprategular apophysis ( Figs 6 View FIGURES 6 – 10 , 16 View FIGURES 13 – 20 , 26 View FIGURES 26 – 29 , 35 View FIGURES 32 – 37 : M, DSA) and not considered an embolic membrane (see Hormiga 1994). The tarsal claws are smooth and not pectinate, the orientation of the flattened inverted J-shaped the paracymbium ( Figs 8 View FIGURES 6 – 10 , 28 View FIGURES 26 – 29 : P) is proximal to distal (longer than wide), a flat triangular process projects from the dorsal surface of the paracymbium ( Figs 8 View FIGURES 6 – 10 , 28 View FIGURES 26 – 29 : P), and the palpal tibial apophysis terminates in a heavily sclerotized refexed barb ( Figs 7, 8 View FIGURES 6 – 10 , 27, 28, 29 View FIGURES 26 – 29 : PTA). Females are further recognized by relatively large clearly visible orangish-yellow spermathecae with posterior margins positioned anterior to the copulatory openings and by the greater width between the ectal margins of the spermathecae than the greatest width of the triangular dorsal plate ( Figs 11 View FIGURES 11 – 12 , 30 View FIGURES 30 – 31 : S, CO, DP).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.