
Cyphophthalmus, JOSEPH, 1868
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2009.00446.x |
|
publication LSID |
lsid:zoobank.org:pub:4CD991A7-6926-4828-863C-029FD0875D1A |
|
persistent identifier |
https://treatment.plazi.org/id/1E1587DD-6A66-FFCE-A3F2-EA7DFD942538 |
|
treatment provided by |
Felipe |
|
scientific name |
Cyphophthalmus |
| status |
|
CYPHOPHTHALMUS JOSEPH, 1868 View in CoL
Revalidated from the synonymy of Siro
Junior synonym: Tranteeva Kratochvíl, 1958: 374 .
Trenteeva: Juberthie, 1970: 1386 ; Juberthie, 1991: 263 (incorrect subsequent spelling).
Diagnosis: Distinct from all other sironids by the specific structure of the coxosternal complex, spermatopositor with a pair of well-developed exterior movable fingers with opposing tips, and presence of two anal glands.
Type species: Cyphophthalmus duricorius Joseph, 1868 by monotypy.
Distribution: The Balkan Peninsula and western Asia Minor.
According to the published data, the genus Cyphophthalmus defined in this way contains 18 species:
Cyphophthalmus duricorius Joseph, 1868 – wide distribution in Slovenia and border regions of Austria, Italy, and Croatia. According to the available data, this species has the largest known distribution area within the genus.
Cyphophthalmus gjorgjevici (Hadži, 1933) – known from a cave near Skopje, Macedonia.
Cyphophthalmus klisurae ( Hadži, 1973) View in CoL – a cave in Rugovska Klisura in Metohia, Serbia.
Cyphophthalmus beschkovi (Mitov, 1994) View in CoL comb. nov. – a cave in western Stara Planina Mountain, Bulgaria.
Cyphophthalmus bithynicus ( Gruber, 1969) View in CoL stat. nov. – Uludağ Mountain near Bursa (north-west Asia Minor, Turkey).
Cyphophthalmus corfuanus ( Kratochvíl, 1938) View in CoL stat. nov. – Corfu, Greece.
Cyphophthalmus yalovensis ( Gruber, 1969) View in CoL stat. nov. – near Yalova in north-west Asia Minor ( Turkey).
Cyphophthalmus eratoae ( Juberthie, 1968) View in CoL – several localities in central and western Greece.
Cyphophthalmus minutus ( Kratochvíl, 1938) View in CoL – a cave on the southern slopes of Orjen Mountain in Konavli (southern Dalmatia, Croatia).
Cyphophthalmus montenegrinus ( Hadži, 1973) View in CoL – a cave near Virpazar ( Montenegro). Despite repeated attempts this peculiar species could not be found in its type locality (cave Velja Spila near Seoce). In this cave and some others in the close vicinity another two species of the genus Cyphophthalmus View in CoL sympatrically appear: Cyphophthalmus cf. teyrovskyi View in CoL and C. cf. minutus View in CoL . Cyphophthalmus cf. teyrovskyi View in CoL may correspond to C. montenegrinus View in CoL by some, primarily morphometric, characteristics; however the absence of the most prominent and distinct characteristics according to Hadži (lateral tooth-like projections on claws IV and the specific ‘inner’ longitudinal and transverse thickenings of the dorsal scutum cuticle) prevents one from drawing definite conclusions about the status of these species. Considering that the type material has been lost this species could be transferred to nomina dubia.
Cyphophthalmus noctiphilus Kratochvíl, 1940 View in CoL – from a cave near Split, Dalmatia.
Cyphophthalmus ohridanus Hadži, 1973 View in CoL – from Ohrid, Macedonia.
Cyphophthalmus serbicus Hadži, 1973 View in CoL – wide distribution in eastern and central Serbia.
Cyphophthalmus silhavyi Kratochvíl, 1938 View in CoL – from a cave in Konavli, southern Dalmatia – Croatia.
Cyphophthalmus teyrovskyi Kratochvíl, 1938 View in CoL – from a cave on the island Mljet, Dalmatia – Croatia.
Cyphophthalmus paradoxus (Kratochvíl, 1958) View in CoL comb. nov. – from caves in western Mount Stara Planina, Bulgaria.
Cyphophthalmus ere Karaman, 2008 View in CoL – distributed in western Serbia ( Karaman, 2008).
Cyphophthalmus nonveilleri Karaman, 2008 View in CoL – from caves in south-western Serbia.
Cyphophthalmus markoi Karaman, 2008 – from Demir Kapija, Macedonia.
The status of the subspecies Siro duricorius bolei Hadži, 1973 from the cave Vilina Jama in Boka Kotorska, Montenegro, is unclear because it appears that the type and only specimen of this species (a female) has not been preserved. In the same cave I collected specimens which belong to the complex of the species C. minutus (troglobite), and in a wider area of the region (the hill Vrmac) there exists an endogean species – Cyphophthalmus martensi sp. nov. (related to the species C. duricorius ). The rather rough description of S. duricorius bolei corresponds to neither of the two species that I found in that area ( Karaman, 1993). The morphometric characteristics given in the description would correspond more to the species C. cf. minutus , but none of the most distinct morphological characteristics of this species are mentioned in the description. It is likely that this taxon also should be transferred to nomina dubia.
Remarks: Results of the long-standing work on collecting and studying Balkan sironids indicate that this group has a rich fauna, with a long and complex genesis. Distinctiveness of characters in relation to other sironids and the numerous taxa often with overlapping distribution areas indicate an independent, long and dynamic history of the genus. According to my own records and P. Mitov’s unpublished data, including described as well as collected but as yet undescribed species, the genus Cyphophthalmus has more than 50 species. The number may further increase, considering that there are still wide areas poorly studied or unexplored as regards this fauna ( Turkey, Bosnia and Herzegovina, Albania etc.) and given the high levels of endemism of the taxa known to date.
Two decades of my more or less intensive work on the collection and taxonomic classification of Balkan Cyphophthalmi (I have had the opportunity to explore in more detail some parts of former Yugoslavia), together with material I have been given to analyse, resulted in a thorough study of around 50 species of the genus Cyphophthalmus . Of these number, more than 30 are new species awaiting formal description. In this paper I decided to describe 12 species that represent the diversity of morphological characters in the genus. This will be enough to illustrate the significance of differences that may seem of minor value for some colleagues in the field. Descriptions of the remaining species will be published at a later time.
The genus Cyphophthalmus has a special and distinct position within the family, owing primarily to the ventral prosomal complex and spermatopositor morphology. However, although the results of molecular analyses clearly indicate a monophyletic origin of the genus, its precise position in the family has not yet been established ( Boyer et al., 2005).
TAXONOMICALLY SIGNIFICANT CHARACTERS OF THE GENUS CYPHOPHTHALMUS
The complex taxonomy of the rich Balkan Cyphophthalmi fauna, with overlapping distribution areas of older and younger troglobitic and endogean species and groups of species (where species sympatrically occur in four cases, two endogean and two in caves), recommend them as a ‘benchmark’ in determining the criteria for the importance of certain taxonomic characters amongst the family.
Conservativeness of morphological characters is a characteristic of Cyphophthalmi, most prominent in the members of the genus Cyphophthalmus . Therefore the study of this genus requires a special approach in determining the criteria of significance of certain taxonomic characters and is actually based on discovering the minute stable differences that will define a species whose members appear more or less distinct by some of their quantitative characteristics and proportions. A series of new valid taxonomic characters was defined in addition to the known ones, of which some have proved significant in determining phylogenetic relationships within the genus.
The male external genital anatomy in opilionids is one of the most valuable differential characteristics of the order and an extremely distinctive one. The morphology of copulatory apparatus shows a certain degree of conservativeness and is therefore a considerably reliable character in phylogenetic studies as well ( Martens, 1976). The application of this character in the study of opilionids started relatively late (1930s) and considerably improved the taxonomy of the whole group. As regards this character and its applicability Cyphophthalmi significantly differ from other opilionids. The discovery of spermatophores in this group ( Karaman, 2005a) has revealed a different role of male external genitals compared to other opilionids. Male genital apparatus in the Cyphophthalmus species has a role of producing spermatophores; therefore the adequate term to designate its external part is spermatopositor (first used for this group by van der Hammen, 1985). Such an apparatus is exposed primarily to the selective pressure of its functionality and efficacy. In opilionids from other suborders (known to date) male external genitals have the role of a penis, conveying sperm into distal parts of a female’s ovipositor by direct and active penetration through the female’s genital opening. Such an apparatus suffers additional selective pressure from relevant female genital structures, functioning like a key and lock. This advanced way of insemination entails co-evolution of the two structures (i.e. male and female genitals), and often also a complex behavioural pattern that accompanies the copulation act, providing and stimulating reproductive isolation. The structure of male genitals is thus specific in many of its details.
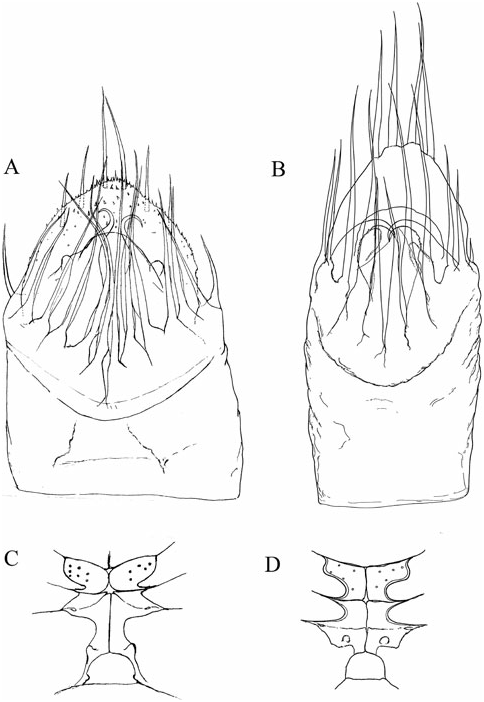
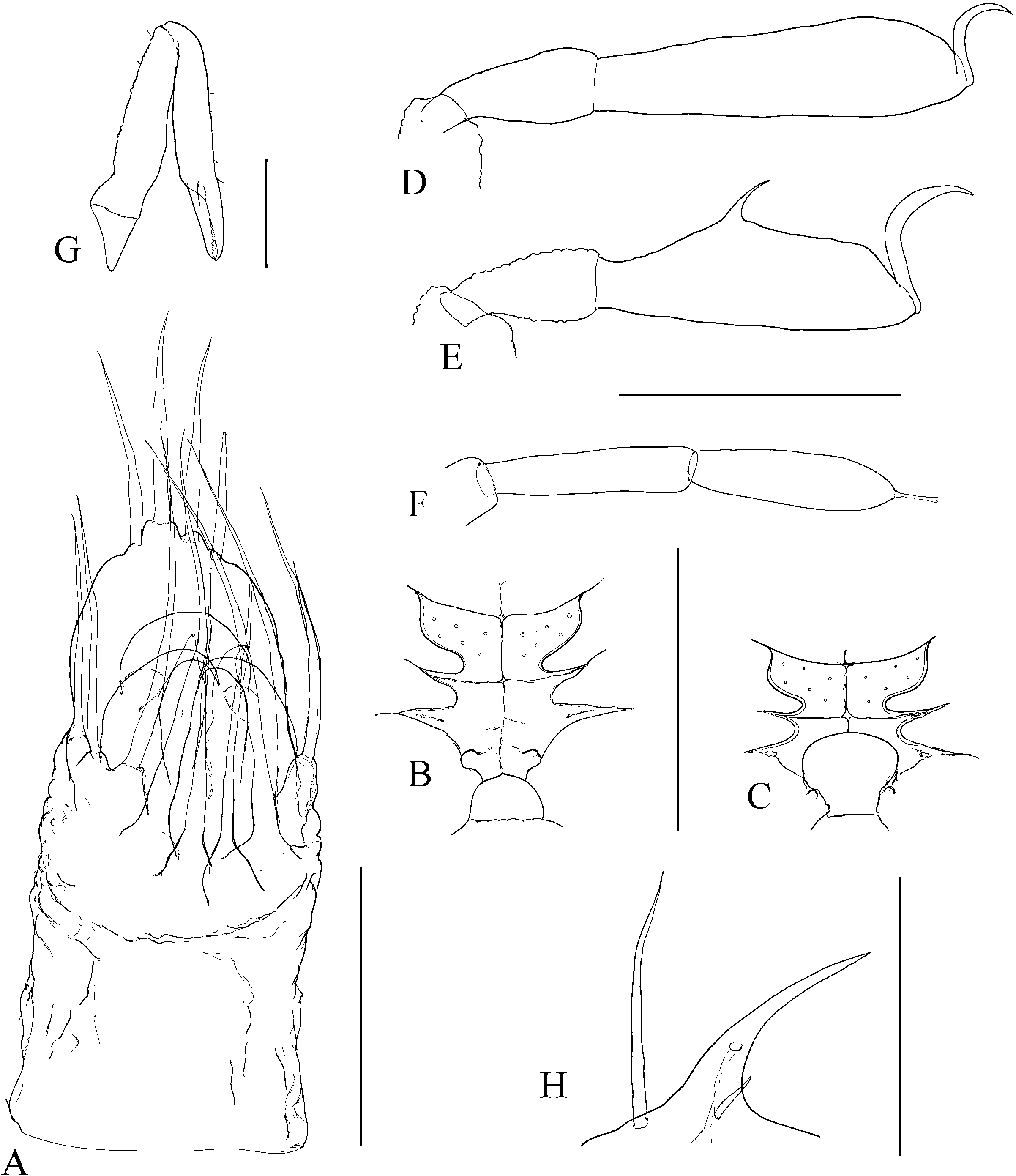
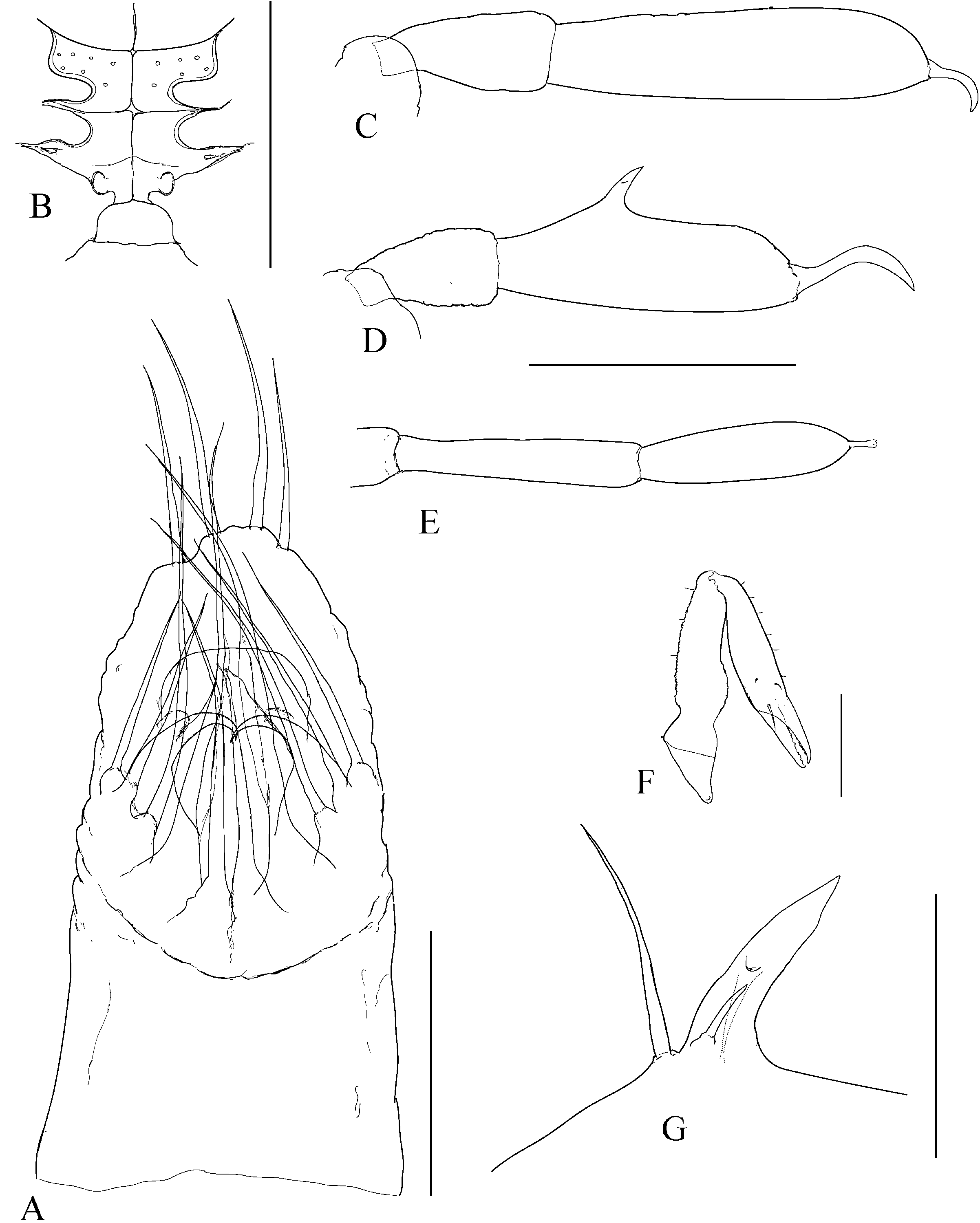
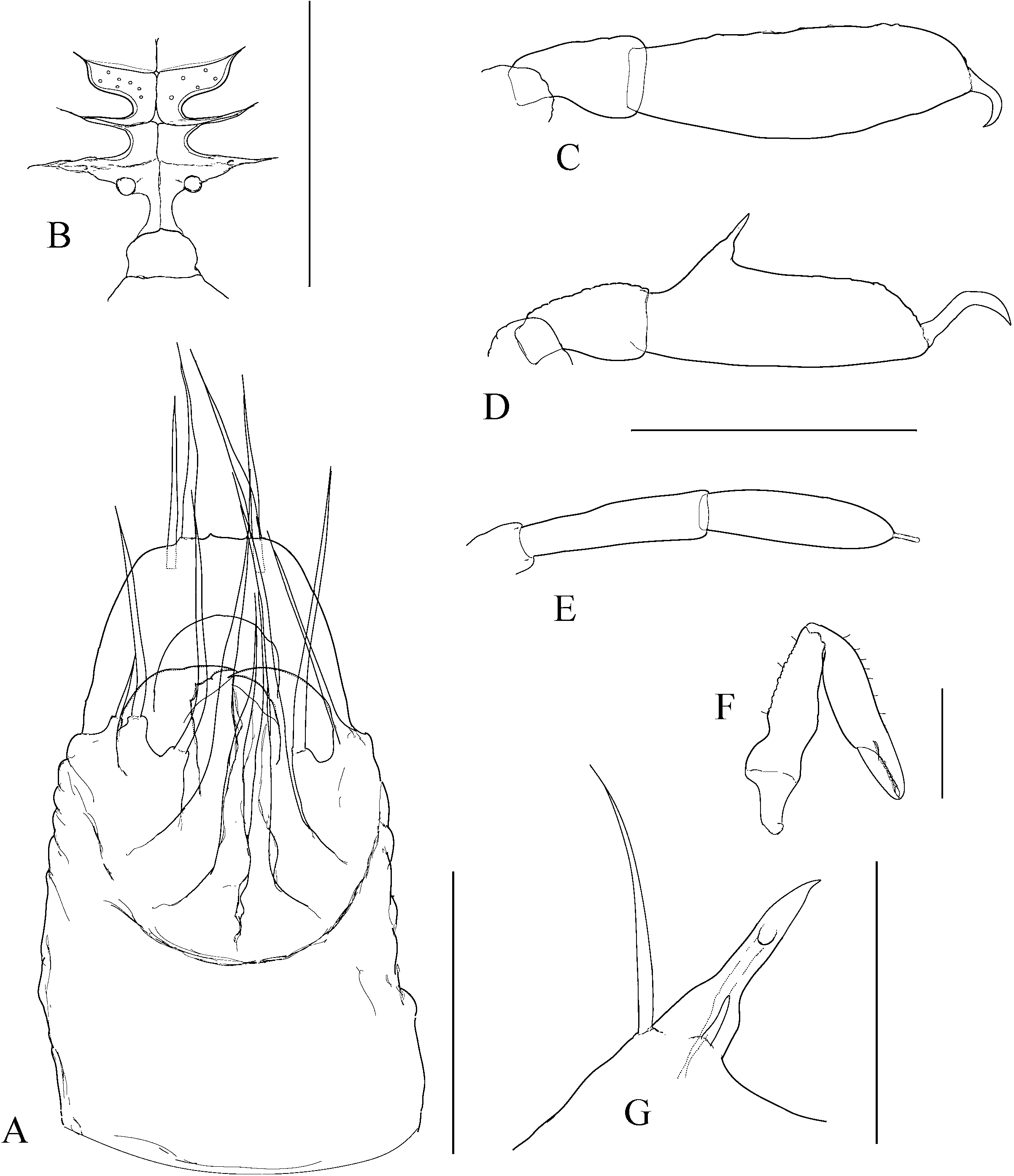
The character of the spermatopositor is the most prominent and distinctive feature of the genus Cyphophthalmus , and a considerably conservative one. This conservativeness is reflected primarily in its qualitative characteristics, and in this sense variations from a typical structure ( Fig. 1C View Figure 1 ) are almost negligible on the level of the whole genus Cyphophthalmus . On the one hand, the conservativeness is reflected in the function of male genitals in this genus. On the other hand, there is some, primarily quantitative, variability which makes it hard to single out valid and taxonomically applicable characteristics of the spermatopositor anatomy. Although habitual differences in the general spermatopositor appearance amongst individual taxa may sometimes appear striking ( Figs 23A View Figure 23 , 29A View Figure 29 ), a more detailed analysis reduces the differences to only a small number of structural characters:
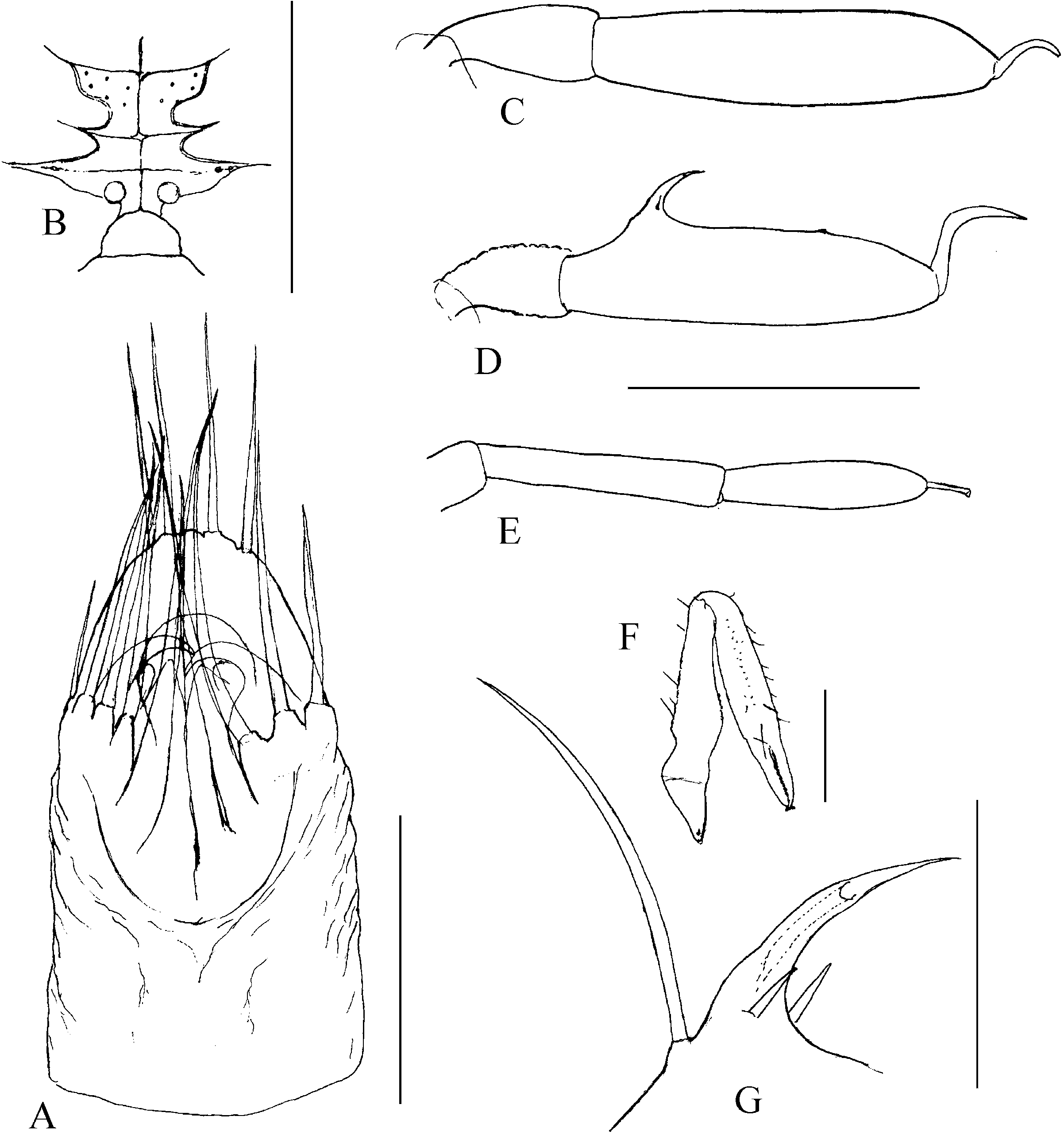
1. Middle movable fingers (digiti mobiles mediales) are considerably reduced compared to other members of the family Sironidae . A similar degree of reduction is usually common to a group of related taxa. There are two basic types of middle movable fingers in a genus: well developed (as long as lateral movable fingers) ( Fig. 2F View Figure 2 ) with tips bent dorsally; or considerably reduced (shorter than lateral movable fingers, sometimes inconspicuous), with anteriorly directed tips ( Fig. 2E View Figure 2 ). The reduced size and soft structure make them almost unnoticeable in some cases. This could explain their ‘absence’ in the species C. eratoae , which its author regarded as the species’ most distinctive characteristic.
2. Lateral movable fingers (digiti mobiles laterales) have the appearance of strong claws ( Fig. 2E, F View Figure 2 ) and are unique to the members of the genus Cyphophthalmus . The tips of these ‘claws’ are curved inwards, opposing one another. The extent of curving and sclerotization, their length, width at the base, and presence or absence of terminal dentations, although variable to a certain degree, indicate species-specific characteristics.
3. The terminal lobe is one of the most valuable taxonomic characters of spermatopositor morphology in the genus Cyphophthalmus . Unfortunately, because of its simple structure this character does not have a variety of forms and often has no differential value unless combined with other characters. As a result of its soft structure it often suffers deformation in the preparationmaking process and thus makes an unreliable character in small series of available specimens. Setae (four) distributed on the tip of the terminal lobe, often with specific distribution and length, add significance to the character of terminal lobe. Setae ventrales (three), positioned ventrally on the terminal lobe, have an inconsistent distribution and sometimes also number, and are therefore an unreliable character.
4. Lateral lobes (lobi laterales) are variable as regards their size and the number of setae they carry (three to five), but in some cases they can be valuable.
5. Trunk (spermatopositor from its base to the transverse fold) is a character which can be defined descriptively as long or short. A long trunk is characteristic for some troglobitic species.
6. Setae dorsales (four) are in some cases useful as taxonomic characters. Basically there are two types: the type with a narrow median pair (narrower to considerably narrower in their basal parts compared to the lateral pair) and the type with a wide median pair (as wide as or wider in their basal parts compared to the lateral pair) ( Figs 7A View Figure 7 , 9A View Figure 9 , 17A View Figure 17 ).
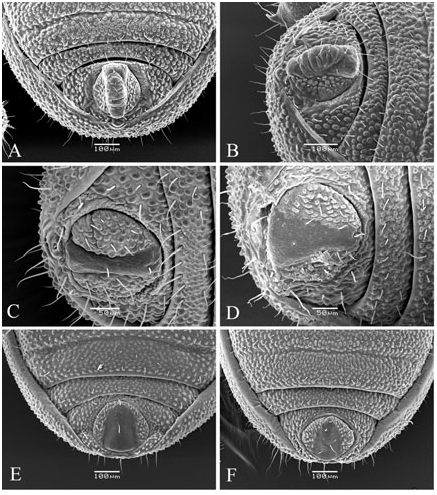
The number of anal gland pores (two) is another character unique to the genus Cyphophthalmus . Distance between the anal gland pores, their orientation, and ornamentation of the posterior opisthosoma are significant in distinguishing phyletic lines within the genus, distinctive in certain phyletic lines and less frequently in a single taxon ( Figs 8E View Figure 8 , 12E View Figure 12 ). Two basic types of the structure can be clearly distinguished in the species of the genus Cyphophthalmus analysed in the present paper. In the Aegean phyletic line ( Figs 34E View Figure 34 , 30E View Figure 30 , 32E View Figure 32 ) the pores are arranged one by the other on the protruded middle portion of the posterior margin of tergite VIII, ventrally orientated, whereas in the Dinaric phyletic line ( Figs 14E View Figure 14 , 24E View Figure 24 , 26E View Figure 26 ) the anal gland pores are wide apart on the posterior margin of tergite VIII, orientated terminally. The studied species from the Gjorgjevici phyletic line are similar to the Dinaric line, the only difference being a smaller distance between the pores ( Fig. 10E View Figure 10 ).
Coxal lobes in males (and to some degree in females) with their specific structure represent another differential character of the genus Cyphophthalmus . The shape and structure of the coxal lobes (ventral prosomal complex) of legs II–IV show taxonomically significant characteristics:
1. Coxal lobes of legs II are a stable character occurring in several forms ( Figs 7B View Figure 7 , 21B View Figure 21 , 31B View Figure 31 ), useful in combination with other characters. This character is equally applicable in males and females. The number and distribution of setae may vary.
2. Coxal lobes of legs III, primarily by their relative length and sometimes by their profile, clearly distinguish some taxa, and are significant in combination with other characters.
3. Coxal lobes of legs IV, by the position of tubercles in the posterior portion and less significantly by their profile, are a clear differential character of certain taxa complexes ( Figs 21B View Figure 21 , 31B View Figure 31 ). (Not of universal significance, however, because in some taxa in which tubercles are not arranged on the anterior margin of the male genital orifice, the position of tubercles and the coxal lobes profile can vary.)
Opisthosomal tip, from a dorsal view, can be rounded or flat to concave. These two shapes can differentiate some phylogenetic lines of the genus Cyphophthalmus .
Lateral profile of dorsum appears in two main shapes: deep (more arched), rounded posteriorly; and shallow (less arched), elongated posteriorly. These two forms generally correspond to some phyletic lines in the genus. Females from the Dinaric phyletic line have as a secondary sex characteristic lateral folds on opisthosomal tergites VII–XI, forming a longitudinal dorsally orientated rim on each side, most prominent on tergite IX (see Karaman, 1993, 2008). On the ventral portion of opisthosoma, opposite the rims, there are ventrolateral distinctly convexly widened sternites of segments IX–XII. The edges of these two structures make dorsal and ventral borders along almost flat surfaces of the lateral sides of that part of opisthosoma. This character, more or less expressed, may be indicative for some species and its presence or absence differentiates the phyletic lines of the genus. This structure may be regarded as a synapomorphy of the Dinaric phyletic line. It is interesting that this characteristic is most prominent in troglobitic members of this line and most reduced in species more adapted to endogean life in ‘arid’ conditions ( Cyphophthalmus martensi sp. nov.). Although the purpose of this prominent characteristic is not fully understood, it is believed to be a hindrance and ballast for animals originating from primarily terricolous ancestors, adapting to life in endogean conditions. If we accept as correct the hypothesis that Cyphophthalmi are primarily terricolous cryptobionts, then the conditions in troglobitic subterranean habitats are closest to the original conditions, which may explain how they retained this characteristic.
The anterior margin of prosoma shows some variability, conspicuous and even peculiar in some cases ( C. conocephalus sp. nov.) ( Fig. 20B View Figure 20 ).
The ozophore orientation and profile can be distinctive. The ozophores are always situated laterodorsally, but their orientation can be lateral ( Fig. 10F View Figure 10 ) to laterodorsal ( Fig. 12F View Figure 12 ). The profile itself can vary from short (length smaller than basal width) to elongated (length greater than basal width) ( Figs 10F View Figure 10 , 12F View Figure 12 ).
The operculum anale can be distinctive in some taxa, despite a certain amount of variability ( Martens, 1978). Differences pertain to the width and height of longitudinal carina in males ( Fig. 3 View Figure 3 ). The carina is extremely well developed in some taxa. This character is one of the most important characters delineating (one of) the three main phylogenetic lines of the genus Cyphophthalmus . The width of the posterior portion of the carina corresponds to the distance between anal gland pores (with rare exceptions), indicating their functional association. This is corroborated by a series of SEM photographs of the posterior body of various Cyphophthalmi species provided in the paper by de Bivort & Giribet (2004). Those photographs clearly show the absence of longitudinal carina of the operculum in species lacking anal gland pores (or having them in unusual positions).
The ovipositor shows certain variation amongst taxa, primarily with regard to the proportions of terminal articles and terminal lobes. Principally, we may differentiate species with stout and with narrow terminal lobes ( Figs 4D View Figure 4 , 5C View Figure 5 ). However, deformation during the slide-making process is possible, so this character is not always fully reliable. Branched apical setae on lobes can be of the type with branches starting from the base of a seta or the type with branches growing from the elongated ‘trunk’ of a seta, including all intermediate types. My studies have shown the most important differences to be in the structure of receptacula seminis ( Figs 4 View Figure 4 , 5 View Figure 5 ). A few main types of the seminal receptacle characterize groups of related taxa and form the basis for defining the three main phyletic lines of the genus. There are three basic types: sigmoid ( Fig. 4A–D View Figure 4 ); saccate with bifurcated bottom ( Fig. 4E–G View Figure 4 ); and simple saccate ( Fig. 5 View Figure 5 ). Generally, there are no significant differences in the structure of seminal receptacles amongst closely related taxa. The distribution and number of simple setae on the terminal lobe of the ovipositor may be distinctive. In most species the terminal lobe is covered with 13 more or less typically distributed setae. The presence of one more seta is characteristic for some species. This seta is positioned mediodorsally at the approximate midpoint of the lobe’s length ( Fig. 4C View Figure 4 ).
Appendages can define a species by their size and proportion, together with other characters. Chelicerae can differ amongst species in article elongation only, i.e. by the relative ratio of the article length and width (they are extremely long in some troglobites, Fig. 23F View Figure 23 ). Pedipalps differ in apotele length in addition to article length. Legs are often clearly distinguishable amongst species by article length and some other specific characteristics ( Figs 23C–D View Figure 23 , 27C–D View Figure 27 ).
The male adenostyle and tarsus IV carrying it are often the most valuable characteristics of appendages, defining a species. The adenostyle appears in two basic shapes, with intermediate forms: spiniform ( Fig. 17G View Figure 17 ), and tubular ( Figs 7G View Figure 7 , 29G View Figure 29 ). The position of the adenostyle on the tarsus also differs amongst species. The tarsal article of leg IV may be characteristic by the relative length and width ratio, as well as by its lateral profile.
The tarsus of leg I may be valuable because of the differences in the length and width ratio, as well as by its lateral profile.
Tarsal claws are generally simple; however, in some taxa (mostly troglobitic) they may have a specific form. The members of the C. ognjenovici subgroup of the C. minutus species-group has enlarged, distinctly falciform claws on legs III, even more prominent on legs IV ( Figs 11E View Figure 11 , 23D View Figure 23 ). In the members of the C. teyrovskyi subgroup these claws are apically dorsoventrally flattened, almost spoon-like.
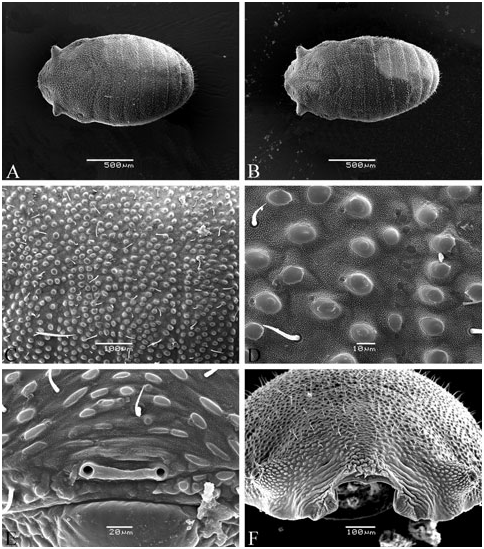
Integument ornamentation can be a very valuable character, often showing the most prominent differences amongst species. However, inclusion of this character in taxonomic analyses requires SEM, because the study of minute details of ornamentation requires great magnification (sometimes more than 1000 times). Differences amongst the species are expressed in the size, shape, distribution, and density of the granules of the first, second, and third rank ( Fig. 6 View Figure 6 ).
The basic criterion for defining a particular Cyphophthalmus species must start from the phylogenetic species concept as the only applicable concept, because the species are almost exclusively found as isolated populations or series of isolated populations. The species are here defined as populations or aggregations of populations with a unique combination of character states. However, the determined criteria that define a particular species may also comply with the biological species concepts that I tend to respect, the fact that I have found in rare so far discovered sympatric populations. Herewith presented characteristics of two relatively close species C. minutus and Cyphophthalmus kratochvili sp. nov., which are sympatric in one cave, are perhaps the best illustration of how some seemingly minor differences in certain characters are indicative in the differentiation and definition of the species of the genus Cyphophthalmus .
The applied criteria have shown to be very stable in geographically very distant populations of rare so far known species of the genus Cyphophthalmus , which have a wider distribution range – C. duricorius and Cyphophthalmus serbicus , as well as in species with somewhat narrower distribution ranges – C. martensi sp. nov. and C. ere .
The presented descriptions are rationalized with a set of unique combinations of character states given in the diagnosis of each species.


















No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Cyphophthalmus
| Karaman, Ivo M. 2009 |
Trenteeva :
| Juberthie C 1991: 263 |