
Idaea josephinae, Guerrero & Hausmann & Ortiz, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.4990.2.10 |
|
publication LSID |
lsid:zoobank.org:pub:F583F46A-35A8-48B1-901A-F01813F15B59 |
|
DOI |
https://doi.org/10.5281/zenodo.5091817 |
|
persistent identifier |
https://treatment.plazi.org/id/2057214B-4A1A-8B65-FF7C-F946FDA114BD |
|
treatment provided by |
Plazi |
|
scientific name |
Idaea josephinae |
| status |
sp. nov. |
Idaea josephinae sp. n.
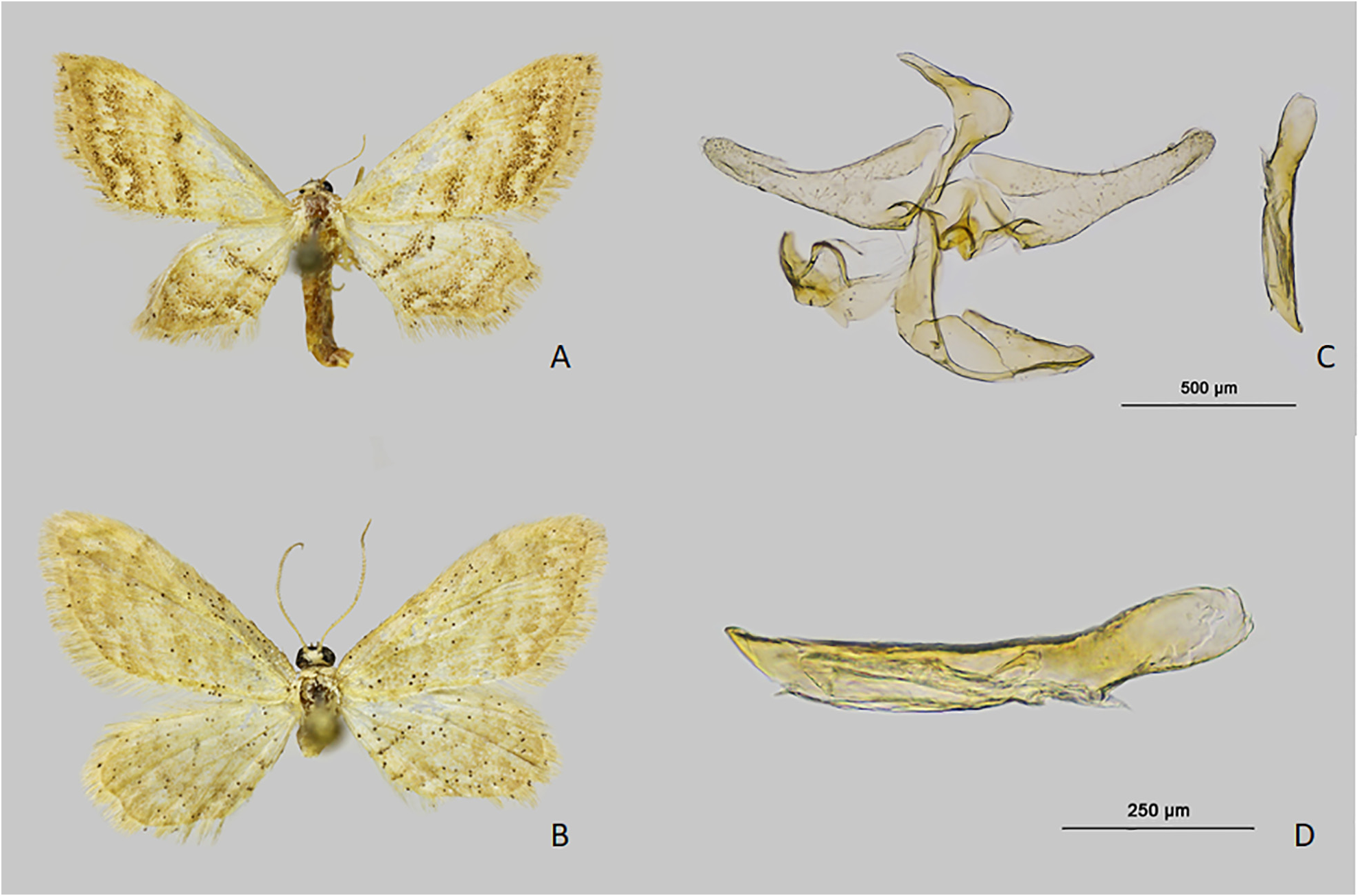
Type material. Holotype. Spain; • 1 ♂; prov. Murcia, Javalí Nuevo, La Contraparada , lat: 37.994746, long: - 1.216436, 60 m; 21 Sep. 2020; leg. J.J. Guerrero; [ Process ID] IBLAO1680-20 ; [genitalia preparation] G 890 ♂ JJ. Guerrero; coll. RCBA ( Fig. 1 A View FIGURE 1 ).
Paratypes. Spain; • 1 ♀; same data as holotype; [ Process ID] IBLAO1706-20 ; coll. ZSM; • 1 ♀; [ Process ID] IBLAO1691-20 ; coll. MNCN ( Fig. 1 B View FIGURE 1 ); • 1 ♀; same data as holotype; 29 Jun. 2020; [ Process ID] IBLAO1690-20 ; coll. RCBA; • 2 ♀♀; prov. Murcia, Las Torres de Cotillas, Rambla Salada , lat: 37.999215, long: -1.246926, 80 m; 8 Jul. 2003; leg. J.J. Guerrero; coll. ZSM ( Fig. 1 C View FIGURE 1 ) ; • 1 ♀, 8 Jul. 2018 [ Process ID] IBLAO1689-20 ; [genitalia prepara- tion] G 891 ♀ JJ. Guerrero ; coll. MNCN ; • 1 ♀, 8 Jul. 2018 [genitalia preparation] G 892 ♀ JJ. Guerrero ; coll. RCBA ; • 1 ♀; prov. Murcia, Las Torres de Cotillas, Embalse de los Rodeos , Río Mula , lat: 38.043687, long: -1.300303, 120 m; 27 Jun. 2003; leg. J.J. Guerrero; [genitalia preparation] G 895 ♀ JJ. Guerrero ; coll. RCBA ; • 1 ♂, 24 Jul. 2003; [geni- talia preparation] G 894 ♀ JJ. Guerrero ; coll. RCBA ; • 2 ♀♀; prov. Murcia, Murcia , Cañada Hermosa, Rambla Salada , lat: 37.9856, long: -1.32313, 150 m; 29 Jun. 2006; leg. J.J. Guerrero; coll. RCBA ; • 2 ♀♀, 17 Jul. 2006; coll. RCBA ; • 1 ♀, 17 Jul. 2006; [ Process ID] IBLAO906-12 ; [genitalia preparation] G 893 ♀ JJ. Guerrero ; coll. RCBA .
Etymology. The species epithet refers to Josephina, first name of the first author’s mother.
Diagnosis. Idaea josephinae belongs to a group of iberian species with one distinct black discal dot in the narrow forewing, slightly concave hindwing margin, creamy white ground colour, and sometimes vaguely marked brownish transverse lines. This species is also similar to the infirmaria species-group including I. saleri , I. tineata , I. infirmaria and l. rhodogrammaria , as outlined in Hausmann (2004). All these species are similar in habitus and share small male genitalia with setose base of uncus, slender and spatulate valva. The cornutus is present in I. infirmaria and I. rhodogrammaria , the truncated or sub-bilobed uncus in I. saleri , I. tineata and I. infirmaria .
Major diagnostic characters are found in the male genitalia ( Figs 2A, B View FIGURE 2 ; 3C, D View FIGURE 3 ). The male genitalia of I. josephinae differ from those of I. tineata by the presence of cornutus and by the presence of an untruncated uncus in I. saleri , narrowed and sharped in I. rhodogrammaria and truncated in I. infirmaria and I. rhodogramaria .
The female genitalia differ from those of I. saleri , I. infirmaria and l. rhodogrammaria by the presence of a broad and bilobed antrum while I. saleri has a very long ductus bursae. The corpus bursae of I. infirmaria and l. rhodogrammaria is sclerotized. A bilobed antrum is present also in I. tineata but the latter differs from I. josephinae by a lateral appendix bursae.
Certain forms of sympatric Idaea fractilineata are very similar, but the spring generation is slightly larger (wingspan ♂ ♀ 12–16 mm), discal spots usually inconspicuous, male hindtarsus shortened, 0.4–0.5 times the length of tibia. Female genitalia smaller, antrum not laterally swollen, ductus bursae shorter and sclerotized. Allopatric North African and Levant Idaea lobaria with slight differences in external appearance, but in male genitalia with a narrower base of the valva and narrower aedeagus without cornutus. Female genitalia similar, with narrower antrum lobes and a slightly longer ductus bursae.
Finally, all species are easily separated by their DNA barcode sequences ( Fig. 4 View FIGURE 4 ).
Description. External characters and abdomen: Small, wingspan ♂ ♀ 10–15 mm (average 13.3 mm; n=11), although the second-generation specimens are much smaller (mean 10.7 mm; n=3). Forewing narrow with slightly pointed apex, hindwing margin slightly concave between M1-M3. Ground colour white or creamy white, sometimes with pale brown suffusion. Transverse lines from pink or salmon-pink to brown, basal, antemedial, medial and postmedial lines sharp, thin and curved towards costa. Subterminal line wider and lobed, terminal line wavy, well defined and wide. Terminal area pale brown with paler wavy line. Cell spots black, sharp, very small, but conspicuous. Antennae of male ciliate-fasciculate, cilia dense and curved. Hindfemur, -tibia and -tarsus slender and of equal length. Fringe scales long, concolorous with transverse lines, sometimes with black basal dots.
Variation. Ground colour white or creamy white, sometimes with pale brown transverse lines with variable dark suffusion. Medial line sometimes conspicuously dark.
Male genitalia. Uncus short, triangular, narrowly truncate at the apex. Gnathos short with a rounded tip. Saccus shortly projecting. Valva shortly spatulate, length 0.9 mm, without spinules at tip. Aedeagus broad, straight, length 0.85 mm. Aedeagus with a narrow, straight, strongly sclerotized cornutus, length 0.2 mm. ( Figs 2A, B View FIGURE 2 ).
Female genitalia. Papillae anales slightly sclerotized and setose. Posterior apophyses twice the length of anterior apophyses. Antrum swollen and laterally bilobed, sclerotized at base. Ductus bursae short and wide. Corpus bursae pyriform, anteriorly wrinkled longitudinally furrowed in its posterior part ( Fig. 2C View FIGURE 2 ).
Distribution and abundance. Iberian endemic restricted to south-easternmost Spain. Rare and local.
Biology. Host plant and early stages are unknown. Bivoltine species, adults have been collected from early June to mid-July and, again in mid-September in xerothermophilous and riparian habitats ( Figs 5A, B View FIGURE 5 ). At a first glance, the species had to be considered as xerothermophilous inhabiting two semi-desert landscapes around seasonal or sporadic water courses at 150 m above sea-level, but later both generations were collected in a lush riverside vegetation area next to the Segura river ( Fig. 5C View FIGURE 5 ). For a detailed description of the xerothermophilous habitat see Guerrero et al. (2018).
Molecular analysis. BIN BOLD:ACD0706 (n=6; Table 1 View TABLE 1 ; sequence length 633 to 658 bp). Intraspecific varia- tion 0%. Judged from COI divergence, the new species is isolated from Idaea lobaria barcodes from Morocco (n=2) and Tunisia (n=1) by genetic distances of 1.9% and 2.1%, the last two clustering to another BIN (BOLD:AAE5597). Levantine populations of I. lobaria from Israel and Jordan (n=5; BOLD:ABZ0361) diverge by 2.8% from Span- ish I. josephinae (cf. Hausmann et al. 2020). Nearest species in Europe: I. consanguiberica (5.7%), I. belemiata (6.1%). At larger genetic distance (7.4%) from I. infirmaria which previously had been suspected as potentially being closely related (cf. Table 1 View TABLE 1 , Fig. 4 View FIGURE 4 ). The new species belongs to a phylogenetically isolated lineage related to the North African and Levantine Idaea lobaria , which is well supported by morphology and genetic data ( Müller et al. 2019; Hausmann et al. 2020).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |