
Platycraninae Brunner v. Wattenwyl, 1893
|
publication ID |
https://doi.org/10.11646/zootaxa.4896.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:0BAD0251-42BC-4C88-BDDC-9622FD1F9F13 |
|
DOI |
https://doi.org/10.5281/zenodo.4384021 |
|
persistent identifier |
https://treatment.plazi.org/id/2570264F-B326-FFAF-98BC-FE36FDF8FAE3 |
|
treatment provided by |
Plazi |
|
scientific name |
Platycraninae Brunner v. Wattenwyl, 1893 |
| status |
|
Platycraninae Brunner v. Wattenwyl, 1893 View in CoL sensu nov.
Type-genus: Platycrana Gray, 1835: 36 View in CoL .
Platycraniae Brunner v. Wattenwyl, 1893: 97 (in part - only the type-genus Platycrana Gray, 1835 View in CoL )
Redtenbacher, 1908: 368 (in part). [ Phibalosomini (II. Sectio Platycraniae]
Platycraniinae Günther, 1953: 557 . [Incorrect spelling based on misspelled Platycrania Westwood, 1859 View in CoL ]
Platycraninae Bradley & Galil, 1977: 190 View in CoL (in part). [Corrected spelling]
Otte & Brock, 2005: 33 (in part).
Acrophyllini Redtenbacher, 1908: 436 (in part).
Cladoxerinae Karny, 1923: 237 (in part).
Lanceocercata Bradler, 2001: 179 (in part).
Pharnaciini Günther, 1953: 555 View in CoL (in part).
Bradley & Galil, 1977: 193 (in part).
Phasminae Karny, 1923: 240 (in part).
Günther, 1953: 554 (in part).
Phasmatinae Bradley & Galil, 1977: 192 View in CoL (in part). [Corrected spelling of Phasminae Karny, 1923: 240 ]
Phryganistriinae Kirby, 1904: 358 (in part).
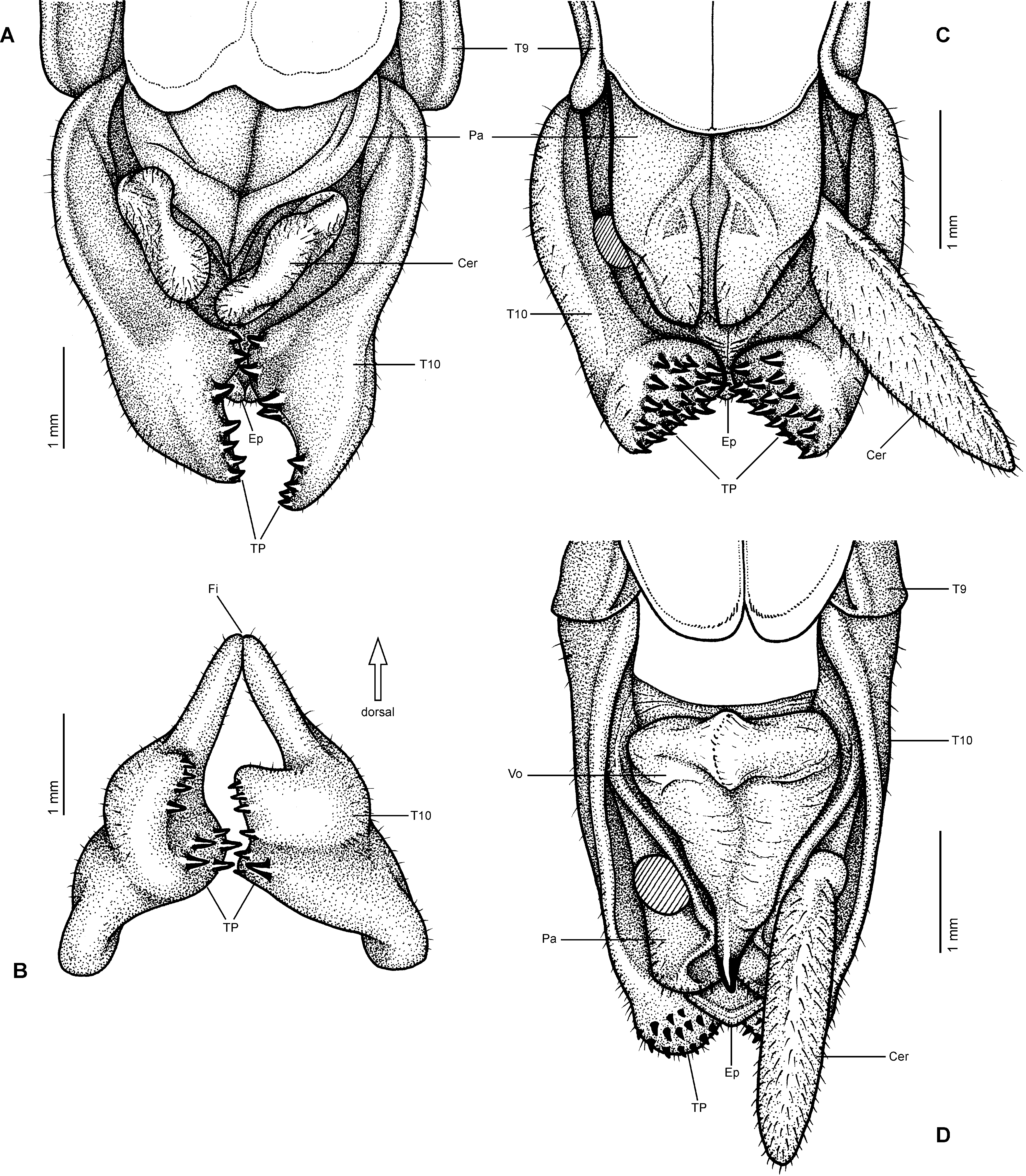
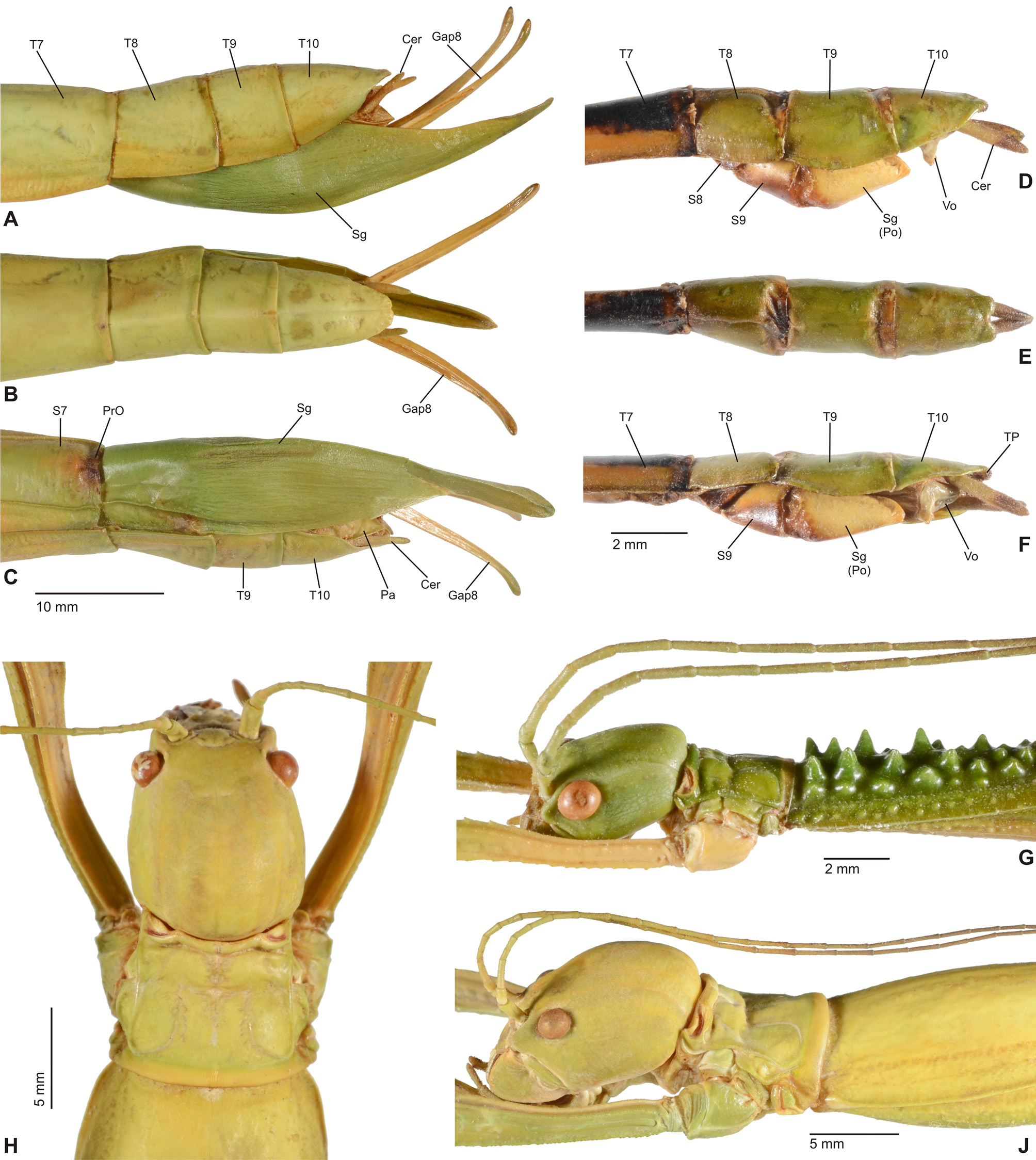
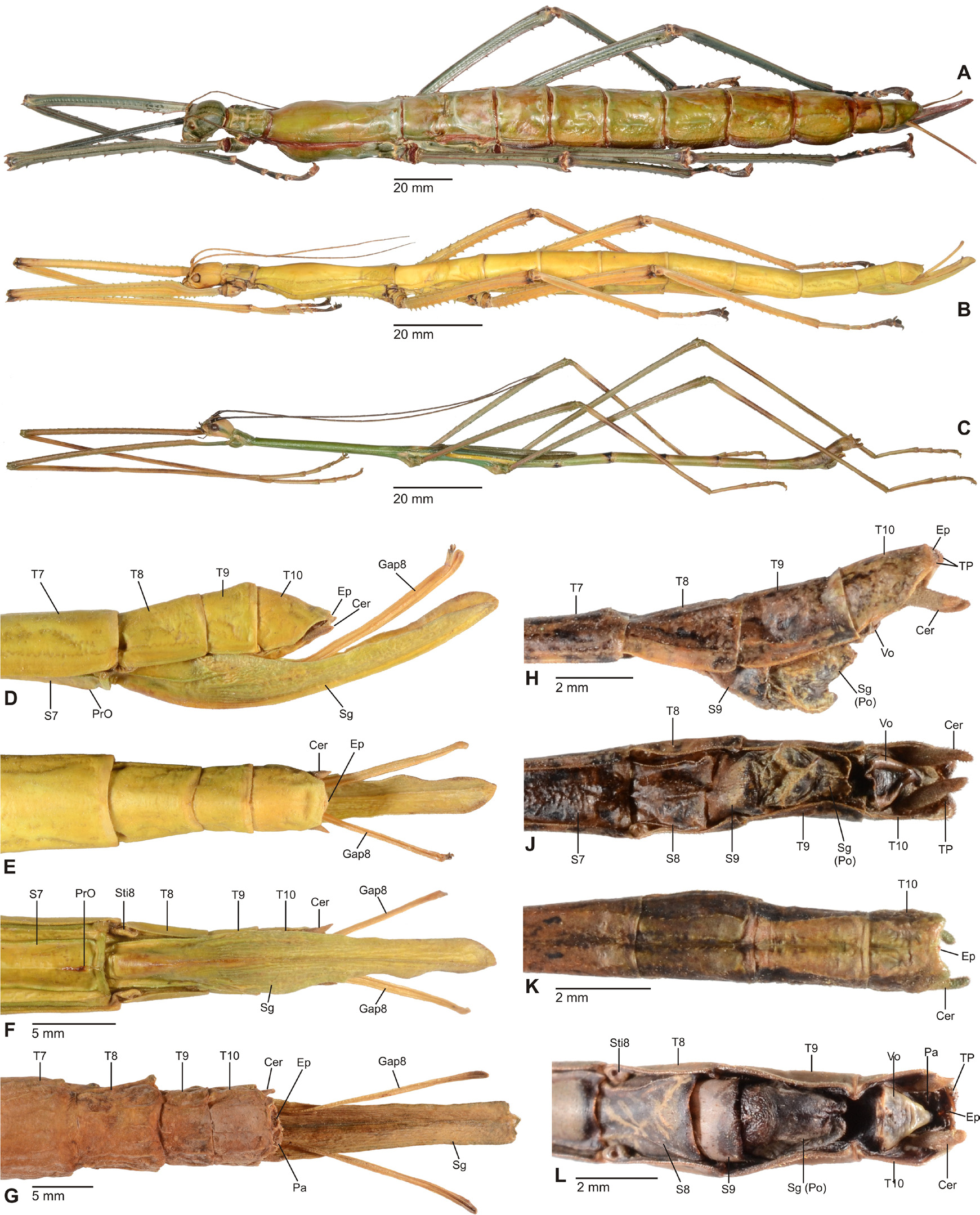
Diagnosis (♂, ♀): Moderately sized to very large Phasmatodea (body length 76.0-300.0 mm) and comprising some of the world’s longest insects. Very strong sexual dimorphism; ♂♂ stick-like, form of ♀♀ ranging from slender to very stocky and sometimes remarkably massive. Both sexes apterous, brachypterous or pterous; apterous or brachypterous ♀♀ often with corresponding ♂♂ brachypterous or with fully developed alae. Body surface ranging from smooth and slightly glossy to moderately sculptured; mesonotum und mesopleurae often tuberculose or spinose. Head ovoid to sub-spherical; no ocelli. No gula, only two small gular sclerites. Antennae much shorter than body; in ♀♀ shorter than in ♂♂. Mesothorax slender in ♂♂, often more or less swollen medially in ♀♀ or either lateral margins of mesonotum or mesopleurae deflexed and/or dentate. Median segment variable in length but mostly shorter than metanotum. Praeopercular organ on sternum VII of ♀♀ indistinct ( Figs. 6C View FIGURE 6 , 11D, 11F View FIGURE 11 ). Terminalia of ♂♂: Anal segment simple and entire ( Figs. 6D View FIGURE 6 , 11H, 11K View FIGURE 11 ), at best slightly tectiform an with posterior margin indented medially; ventral surface of posterior margin with paired, ventral directed thorn-pads (TP, Figs. 6D View FIGURE 6 , 10F View FIGURE 10 , 11J, 11L View FIGURE 11 ). External vomer well developed and sclerotised; mostly triangular in shape and with a single terminal hook ( Figs. 6D View FIGURE 6 , 10F View FIGURE 10 , 11J, 11L View FIGURE 11 ). Terminalia of ♀♀: Gonapophysis VIII strongly elongated, filiform and projecting considerably beyond apex of abdomen ( Figs. 10 View FIGURE 10 A-C, 11D-G); much longer than gonapophysis IX or gonoplacs. No gonangulum present. Subgenital plate prominently keeled longitudinally and usually projecting considerably beyond apex of abdomen; shape variable ( Figs. 10 View FIGURE 10 A-C, 11D-G). Extremities with most or all carinae more or less strongly dentate, serrate and/or occasionally with single lobes. Profemora trapezoidal in cross-section, compressed and curved basally; the medioventral carina indistinct and roughly midways on ventral surface of profemur; the posterodorsal carina moderately raised. Meso- and metafemora trapezoidal in cross-section. Tibiae without an area apicalis. Basitarsus longer than the following two segments combined.
Eggs ( Figs. 12 View FIGURE 12 J–M): Small to large, shape variable; ranging from simply ovoid over almost spherical to considerably longer than high with the cross-section distinctly oval and capsule surrounded by a dorso-ventral bulge. Surface of capsule ranging from smooth and glossy to strongly sculptured. Internal micropylar plate closed, no median line. Operculum flat, capitulum present but not stalked.
Comments: The subfamily Platycraninae was based on the genus Platycrana Gray, 1835 . As discussed in detail above, the type-genus Platycrana is not closely related with all other genera that were traditionally attributed to the subfamily. These genera are here removed from Platycraninae and accommodated in Megacraniinae subfam. nov.. As a result, Platycrana is the only genus that remains in Platycraninae sensu nov.. Close relation between Platycraninae sensu novo and the tribe Stephanacridini Günther, 1953 has already been suggested by Hennemann & Conle (2008: 57) and based on a prolific set of morphological characters this relation is supported herein. Common characters include the lack of a gula (only two small gular sclerites present), small and more or less cylindrical cerci of both sexes, a simple and entire anal segment ( Figs. 10D View FIGURE 10 , 11K View FIGURE 11 ) and well developed external vomer of ♂♂ ( Figs. 6D View FIGURE 6 , 10 View FIGURE 10 E-F, 11J, 11L), as well as the more or less prominently elongated subgenital plate and often enormously elongated and filiform gonapophysis VIII of ♀♀, that considerably project beyond the apex of the abdomen ( Figs. 10 View FIGURE 10 A-C, 11D-G). The eggs share a flattened operculum and knob-like central capitulum ( Figs. 12 View FIGURE 12 J-M). All these features are not only shared by Platycrana and Stephanacridini , but they also distinguish both clades from Megacraniinae subfam. nov. and the entire family Phasmatidae s. str. (= Lanceocercata). Consequently, Stephanacridini was removed from Phasmatidae s. str. (= Lanceocercata) by Hennemann & Conle (2008) which has received strong support by subsequent phylogenetic approaches based on morphological data (Buckley et al., 2009) and molecular data (Buckley et al., 2009; Robertson et al., 2018). These studies have moreover recovered Stephanacridini as the sister group of Phasmatidae s. str. (= Lanceocercata) but Stephanacridini has since remained without a subfamiliar affiliation. The here revealed close relation to Platycrana places Stephanacridini as a subordinate taxon of Platycraninae sensu nov., which consequently means that automatically Platycraninae , instead of Stephanacridini , should now be considered as the possible sister taxon of Phasmatidae s. str. (= Lanceocercata). Platycraninae senso nov. comprises two tribes, i.e. the monotypical Platycranini , which only contains its type-genus Platycrana , and Stephanacridini .
Simon et al. (2019) have obtained Xenophasmina Uvarov, 1940 , an Indo-Chinese genus that currently is considered to be a member of the polyphyletic subfamily Xeroderinae , as the sister taxon of Phasmatidae s. str. (= Lanceocercata). Moreover, these authors considered that Xenophasmina might be a member of Stephanacridini . From a morphological point of view however, there is absolutely no support for this assumption. Although ♂♂ of Xenophasmina possess an entire anal segment and a well developed vomer, both sexes have notably enlarged, dorsoventrally flattened and more or less foliaceous cerci, that remarkably resemble the cerci found throughout Phasmatidae s. str. (= Lanceocercata) and ♀♀ lack the elongated and filiform gonapophysis VIII seen iStephanacridini. Moreover, the eggs of Xenophasmina lack the knob-like central capitulum seen in Stephanacridini but have a strongly raised opercular structure with a central hollow instead and the internal micropylar plate is open with a distinct median line (closed in Stephanacridini ). Other remarkable morphological characters, such as the notably flattened head, laterally lobed abdominal terga VII-X of both sexes and notably deflexed, undulate and strongly setose lower outer carinae of all legs in ♀♀ distinguish Xenophasmina from Stephanacridini . Hence, an inclusion of Xenophasmina in future analyses appears desirable in order to uncover the relationships of the genus.
Distributuion: Taiwan, Peninsular Malaysia, Borneo, Philippines, Wallacea, New Guinea, Solomon Islands, NE-Australia, Micronesia and Melanesia ( Fiji, Tonga, Samoa, Vanuatu, New Caledonia & Western French Polynesia).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Platycraninae Brunner v. Wattenwyl, 1893
| Hennemann, Frank H. 2020 |
Platycraninae
| Bradley, J. C. & Galil, B. S. 1977: 190 |
Phasmatinae
| Bradley, J. C. & Galil, B. S. 1977: 192 |
| Karny, H. H. 1923: 240 |
Pharnaciini Günther, 1953: 555
| Gunther, K. 1953: 555 |
Cladoxerinae
| Karny, H. H. 1923: 237 |
Phasminae
| Karny, H. H. 1923: 240 |
Acrophyllini
| Redtenbacher, J. 1908: 436 |