
Squalus acanthias Linnaeus, 1758
|
publication ID |
https://doi.org/10.11646/zootaxa.4133.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:E7276A63-67C8-4BC5-8419-2EBDAE4432B0 |
|
DOI |
https://doi.org/10.5281/zenodo.6075719 |
|
persistent identifier |
https://treatment.plazi.org/id/282F878E-FF8E-FF91-14C0-244EFE1A5FFD |
|
treatment provided by |
Plazi |
|
scientific name |
Squalus acanthias Linnaeus, 1758 |
| status |
|
Squalus acanthias Linnaeus, 1758 View in CoL
( Figs. 2–15 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 , Tables 1–4 View TABLE 1 View TABLE 2 View TABLE 3 View TABLE 4. E )
Spotted spiny dogfish; Piked dogfish; Cação-bagre-espinhoso (Portuguese)
Squalus acanthias Linnaeus, 1758: 233 View in CoL View Cited Treatment (original description, “Oceano Europaeo”); Rafinesque, 1810: 45 (listed; Sicily); Gill, 1862: 405 (cited; global); Poey, 1868: 213, 454 (cited; Cuba); Berg, 1895: 5, 6 (listed; Argentina, Uruguay); Jordan & Evermann, 1896: 54 (listed; North and Central America); Schreiner & Ribeiro, 1903: 79 (listed; Brazil); Regan, 1908: 45, 46 (identification key, listed; global); Garman, 1913: 192, plates 14 (figs. 1–4), 43 (figs. 9, 10), 59 (figs. 1, 2) (description; global); Fowler, 1936: 69 –71, figs. 19, 20 (revision; Eastern Atlantic); Bigelow & Schroeder, 1948: 455 –473, figs. 87 (A– D), 88 (revision; Northwestern Atlantic); Bigelow & Schroeder, 1953: 47–51, fig. 17 (description, cited; Western North Atlantic); Bigelow, Schroeder & Springer, 1953: 221 (cited; Western Atlantic); Bigelow & Schroeder, 1957: 30, fig. 3D (description; global); Garrick, 1960: 520, figs. 1 (A–C), 3 (G–M), 5 (revision; New Zealand); Bass et al., 1976: 13, 14, figs. 8 (F–G), 9 (revision; Eastern South Africa); Cadenat & Blache, 1981: 46 –48; fig. 28 (A–E) (revision; Mediterranean Sea); Lucena & Lucena, 1981: 2, fig. 3 (listed; Brazil); Compagno, 1984: 109 -113 (revision; global); Kondyurin & Myagkov, 1984: 118 –120, fig. 1A (revision; Western Atlantic); Menni et al., 1984: 62, 83, 84 (listed; Argentina, Uruguay); Myagkov & Kondyurin, 1986: 1 –20, fig. 1 (A, E, F, H) (revision; Atlantic); López et al., 1996: 7, 8 (listed; Argentina); Cousseau & Perrotta, 1998: 34–35 (description; Argentina); Lessa et al., 1999: 26, 61, 150 (cited, listed; Brazil); Mazzoleni & Schwingel, 1999: 114 (listed; Itajaí, Brazil); Gadig, 2001: 29, 36, 54–57, fig. 27 (description; Brazil); Soto, 2001: 94, 95 (listed; Brazil); Compagno, 2002: 380, 381, 383 (revision; South Atlantic); Nion et al., 2002: 4 (listed; Uruguay); Haimovici et al., 2003: 38, 39 (cited; Brazil); Meneses & Paesch, 2003: 7, 25, 45 (cited; Argentina); Smith & Heemstra, 2003: 61, 62, fig. 5.24 (identification key; description; South Africa); Heemstra & Heemstra, 2004 (cited, Southern Africa): 54; Soto & Mincarone, 2004: 73, 74 (listed; Brazil); Lamilla & Bustamante, 2005: 9, 26 (cited; Chile); Nelson, 2006: 66 (listed; global); Menni & Lucifora, 2007: 3 (cited; Argentina, Uruguay); Pon & Gandini, 2007 (cited; Argentina); Carrier et al., 2010: 44, 127, 139 (cited); Gomes et al., 2010: 44, 45 (cited; Brazil); Menni et al., 2010 (cited; Southwestern Atlantic); Saéz et al., 2010: 623 (identification key; Chile); Viana, 2011: 28 –56 (description; Southwestern Atlantic); Menezes, 2011: 4 (listed; Southern Brazil); Rosa & Gadig, 2014: 92 (listed; Brazil).
Squalus fernandinus Molina, 1782: 188 View in CoL , 189, 285 (original description, not illustrated; Chile); Regan, 1908: 45, 46 (cited; Chile); Bigelow, Schroeder & Springer, 1953: 220–222 (cited; Western Atlantic).
Acanthorhinus acanthias: Bigelow & Schroeder, 1948: 452 (in synonymy of S. acanthias ).
Spinax acanthias: Cuvier, 1817: 130 (cited); Cuvier, 1863: 320 (listed); Bigelow & Schroeder, 1948: 452 (in synonymy of S. acanthias ).
Acanthias vulgaris Risso, 1826: 13 (original description, not illustrated; Mediterranean, Sea); Müller & Henle, 1841: 83 (description); Duméril, 1865: 437 (description); Günther, 1866: 384, 396 (listed; Central America); Macleay, 1881: 366 (description; Australia); Vaillant, 1888: 5 (listed; Argentina).
Acanthias americanus Storer, 1846: 506 (original description; United States of America); Gill, 1861: 60 (listed; North America).
Spinax acantheus: Cuvier, 1863: 320 (cited).
Acanthias lebruni Vaillant, 1888: 5 View in CoL , 13, 14, plate 1, fig. 2 (original description; Chile).
Squalus lebruni: Berg, 1895: 6 View in CoL (description; Argentina, Uruguay); Menni et al., 1984: 62, 84 (listed; Argentina, Uruguay).
Squalus blainvillei: Schreiner & Ribeiro, 1903: 79 View in CoL (listed; Brazil); Miranda Ribeiro, 1907 (in part): 168 (description; Brazil).
Squalus barbouri Howell-Rivero, 1936: 47 View in CoL , 48 (original description, not illustrated; Cuba).
Squalus tasmaniensis Howell-Rivero, 1936 View in CoL (original description, not illustrated; Tasmania); Last et al., 2007: 109–113, figs. 1B, 2B (revision; Tasmania).
Flakeus tasmaniensis: Whitley, 1940: 139 , fig. 150 (listed; Australia).
Koinga whitleyi: Whitley, 1940: 139 , figs. 151, 152 (listed; Australia).
Koinga kirki: Whitley, 1940: 140 , fig. 153 (listed; Australia).
Squalus kirki Phillipps, 1931 View in CoL (original description, not illustrated; New Zealand).
Squalus View in CoL sp. of acanthias group: Figueiredo, 1981: 17 (listed; Brazil); Gomes et al., 1997: 93 (description; Brazil); Marques, 1999 (cited; Brazil).
Syntype. NRM 85, juvenile female, 177 mm TL, unknown locality. Donation by King Gustav IV Adolf of Sweden. Other possible syntypes in Uppsala (refuted by Wheeler, 1991): UUZM 159, juvenile male, 346 mm TL; UUZM 160, neonate female, 380 mm TL. Type locality: “Oceano Europaeo”. Possibly from off Sweden, Finland or Norway.
Material examined (59 SWAO specimens). AMNH 4099, neonate male, 230 mm TL, between the mouth of the rivers Coyle and Gallegos, Santa Cruz, Argentina; BMNH 1936.8.26.17, adult male, 635 mm TL, near Strait of Magellan, Argentina, Southwestern Atlantic Ocean, 52.18°S, 68°W; BMNH 1999.5.4.4, juvenile male, 550 mm TL, Falkland Islands, Southwest Atlantic Ocean; BMNH 1999.5.4.13, juvenile female, 448 mm TL, Falkland Islands; BMNH 1999.5.4.15, adult female, 535 mm TL, Falkland Islands; BMNH 1999.5.4.16, juvenile male, 496 mm TL, Falkland Islands; HUMZ 30173, adult male, unknown TL, off Patagonia, Argentina, 47°S, 6516’W (dissected); HUMZ 30178, adult male, 693 mm TL, off Patagonia, Argentina, 47°S, 6516’W; HUMZ 30200, adult male, 655 mm TL, off Patagonia, Argentina, 47°S, 6516’W; HUMZ 30285, adult male, 720 mm TL, off Patagonia, 47°S, 6516’W; HUMZ 30291, adult male, 595 mm TL, off Patagonia, Argentina, 47°S, 6516’W; HUMZ 30303, juvenile female, 487 mm TL, off Patagonia, 47°S, 6516’W; HUMZ 30310, juvenile female, 520 mm TL, off Patagonia, Argentina, 47°S, 6516’W; HUMZ 30324, adult male, 760 mm TL, off Patagonia, 47°S, 6516’W; HUMZ 107285, juvenile female, 340 mm TL, off Argentina, 4659.5’S, 6516’W (dissected); MCT 7439, adult female, 587 mm TL, Tramandaí, Rio Grande do Sul, Brazil; MCZ 1435-S, neonate male, 175 mm TL; neonate male, 205 mm TL, Suriname; MNRJ 509, juvenile male, 403 mm TL, Rasa Island, Rio de Janeiro, Brazil; MNRJ 513, juvenile female, 440 mm TL, Rasa Island, Rio de Janeiro, Brazil; NMW 83924, neonate male, 215 mm TL, unspecified locality, Brazil; ZMH 104416, adult male, 725 mm TL; adult female, 755 mm TL, near Peninsula del Valdes, Argentina, 5948'W, 438'S; ZMH 104461, two juvenile females, 510, 557 mm TL; one adult female, 742 mm TL, near Peninsula del Valdes, Argentina, 608'W, 433'S; ZMH 104519, two adult females, 670, 745 mm TL, near Cabo Blanco, Argentina, 6112'W, 478'S; ZMH 104951, adult male, 715 mm TL, near Santa Cruz, Argentina, 6340'W, 490'S; ZMH 104955, seven neonates males, 187–192 mm TL; five neonate females, 185–200 mm TL, near Santa Cruz, Argentina, 6546'W, 4750'S; ZMH 104968, six juvenile male, 226–233 mm TL; four juvenile female, 225–235 mm TL, near Cabo Blanco, Argentina, 6518'W, 470'S; ZMH 107911, four juvenile males, 213– 225 mm TL; six juvenile females, 215–223 mm TL, near Santa Cruz, Argentina, 640'W, 460'S; ZMH 108038, adult male, 670 mm TL, near Cabo Blanco, Argentina, 650'W, 470'S; ZMH 115469, juvenile male, 322 mm TL, near Santa Cruz, Argentina, 6324'W, 4551'S.
Diagnosis. Squalus acanthias from the Southwestern Atlantic Ocean is distinguished from all local congeners by the following unique characters: rounded white spots dorsolaterally on body; anterior margin of nostrils unilobate; origin of first dorsal-fin spine behind the free rear tips of the pectoral fins; first dorsal-fin spine length much lower than fin height (first dorsal-fin spine length 1.5%–2.8% TL vs. first dorsal-fin height 6.0%–8.1% TL); second dorsal-fin spine very elongated (1.9%–4.5% TL), taller than second dorsal-fin apex; interdorsal distance 1.8 (1.5–2.2) times greater than dorsal-caudal distance; pectoral-pelvic distance 1.0 (0.6–1.3) times pelvic-caudal distance; dermal denticles unicuspid; coracoid bar with segmented processes anterolaterally.
Description. External morphology. Measurements and counts are summarized in Tables 1–2 View TABLE 1 View TABLE 2 (in the description of S. acanthias below, single values represent the mean followed by the range for non-type specimens from which data were taken). Body fusiform and slender, slightly more arched anteriorly as of posterior margin of spiracle to level of pectoral fin insertion; head height 0.8–1.1 times trunk height, and 0.8–1.5 times abdomen height. Head flattened, small (its length 18.4%–23.3% TL) and broad, its width 1.2 (1.0–1.5) times trunk width and 1.4 (1.1–2.2) times abdomen width. Snout somewhat elongate (5.8%–8.1% TL), slightly obtuse in ventral view. Nasal apertures located laterally, small and slightly oblique; anterior margin of nostril unilobate or slightly bilobate in young specimens ( Fig. 3 View FIGURE 3 ); distance from snout tip to nostrils 1.0 (0.9–1.2) times distance from nostrils to upper labial furrow; prenarial length corresponding to half of preoral length; internarial distance 0.9 (0.6–1.4) times eye length. Eyes oval with anterior margin rounded and posterior margin angular, not notched; length 2.0 (1.4–3.5) times eye height; eyes closer to snout tip than to first branchial arch. Prespiracular length 1.7 (1.5–2.0) times preorbital length and 0.6 (0.5–0.6) times pre-pectoral length. Spiracles crescent-shaped, located posterodorsally to eye, oblique in relation to longitudinal axis of body; diameter at least one-quarter eye length. Prebranchial length 1.5 (1.3–1.7) times prespiracular length. Gill slits concave, located in front of pectoral fins, tall with fifth gill slit 1.2 (0.7–1.7) greater in height than first gill slit.
Measurements Holotypes Squalus S. barbouri S. tasmaniensis MCZ 1463-S MCZ 146-S N Range x SD TL (mm) 267.0 245.0 21 200.0–880.0 487.7 212.3 PCL 77.2 77.6 21 31.2–81.1 76.3 10.4 PD2 59.2 57.1 21 53.4–62.2 58.8 2.1 PD1 34.5 30.6 21 29.1–33.7 32.4 0.9 SVL 52.4 49.0 21 44.2–52.8 49.7 2.3 PP2 54.3 44.9 21 41.9–50.3 46.8 2.1 PP1 21.8 19.6 21 17.8–22.8 20.4 1.2 HDL 22.4 19.9 21 18.4–23.3 20.9 1.3 PG1 18.5 16.4 21 15.5–20.5 17.4 1.2 PSP 12.6 9.8 21 9.9–14.1 11.3 1.2 POB 7.0 5.4 21 5.8–8.1 6.9 0.6 PRN 5.1 3.3 21 3.8–5.1 4.5 0.4 POR 9.9 7.6 21 7.8–10.9 9.1 1.0 INLF 4.9 3.6 21 3.7–5.6 4.4 0.5 MOW 6.6 5.6 21 6.5–8.1 7.2 0.4 ULA 2.5 2.2 21 2.0–2.9 2.3 0.2 INW 2.8 3.2 21 2.8–4.8 3.4 0.4 INO 7.2 6.2 21 6.5–9.7 7.4 1.0 EYL 3.5 3.1 21 2.7–5.7 3.7 0.9 EYH 0.9 1.6 21 1.4–3.6 1.9 0.5 SPL 1.3 1.8 21 0.9–2.1 1.4 0.4 GS1 1.7 1.0 21 1.3–2.3 1.7 0.3 GS5 1.8 1.6 21 1.2–2.9 2.0 0.3 IDS 18.9 19.9 21 17.1–22.7 20.2 1.7 DCS 10.8 11.9 21 9.6–11.9 11.1 0.6 PPS 24.9 22.2 21 16.5–26.8 22.8 2.5 PCA 22.2 22.4 21 20.3–28.1 22.8 1.9 D1L 12.1 13.2 21 10.5–13.1 12.1 0.7 D1A 11.5 11.2 21 8.5–11.6 9.5 0.8 D1B 7.0 7.9 21 6.4–7.5 7.0 0.3 D1H 7.8 6.9 21 6.0–8.1 6.7 0.4 D1I 5.7 5.8 21 4.4–6.0 5.2 0.4
Mouth arched and evidently broad, its width 1.6 (1.4–1.9) times prenarial length and 2.1 (1.6–2.5) times internarial space; upper labial furrow slender and large, its length corresponding to 2.0%–2.9% TL; lower labial furrow inconspicuous. Teeth unicuspid, similar in both jaws, flattened labial-lingually and laterally overlapped; teeth broad, although short at crown; upper teeth smaller than lower teeth; cusp thick and elongate, oblique and directed laterally (cusp strongly vertical in most adult males); convex mesial cutting edge; both distal and mesial heels rounded; apron well elongated in both jaws; one intermediate tooth present at least in upper jaw, usually smaller than the following teeth, with cusp completely vertical, short apron, and both distal and mesial heels markedly rounded; two series of functional teeth in upper and lower jaws; teeth rows varying from 14– 1–14 in upper jaw and 11– 1–12 in lower jaw ( Fig. 4 View FIGURE 4 ).Origin of first dorsal fin posterior to free rear tips of pectoral fin; in young specimens, origin over free rear pectoral tips (pre-first dorsal length 29.1%–33.7% TL in neonates and juveniles); horizontal distance between origins of pectoral fin and first dorsal fin 1.8 (1.2–2.3) times preorbital length. First dorsal fin vertical and markedly wide at its fin web, relatively large (10.5%–13.1% TL) and low, its length 1.8 (1.5–2.1) times its height, its height and base length 1.0 (0.9–1.2) times preorbital length; anterior margin convex, posterior margin moderately concave near free rear tip; apex rounded, and free rear tips pointed; inner margin length 0.8 (0.6–0.9) times fin height. Origin of first dorsal-fin spine slightly posterior to free rear tips of pectoral fins. First dorsal-fin spine conspicuously slender and low, its length not greater than half dorsal-fin height, never reaching fin apex. Interdorsal distance 1.0 (0.8–1.3) times prepectoral length and 1.8 (1.5–2.2) times dorsal-caudal distance. Pre-second dorsal length 4.3 (3.7–5.2) times the anterior margin length of pectoral fin and 2.9 (2.6–3.2) times dorsal caudal lobe length. Second dorsal fin rather oblique and broad at its fin web, relatively large, its length 1.0 (0.9–1.2) times first dorsal-fin length, and 2.5 (1.9–3.0) times second dorsal-fin height; anterior margin convex and posterior margin falcate; apex rounded, and free rear tip pointed; inner margin elongate, its length 1.0 (0.8–1.1) times fin height. Second dorsal-fin spine slightly inclined, elongated (1.9%–4.5% TL) and reaching the fin apex (in some adult specimens, second spine exceeds the fin apex); slender and low, its length 0.7 (0.4–0.9) times second dorsal-fin height and 1.8 (1.3–2.5) times first dorsal-fin spine length; second dorsal-fin spine broader than first dorsal-fin spine, its base length 1.3 (1.0–1.5) times first dorsal-fin spine base length.
Pre-pectoral length 0.6 (0.6–0.7) times pre-first dorsal length, 0.4 (0.4–0.5) times pre-vent length. Pectoral fins with both anterior and inner margins convex, and posterior margin straight; anterior margin length 1.6 (1.4–1.6) times posterior margin length, but its tip reaches same level of its apex; apex and free rear tips evidently rounded, but not lobe-like; pectoral fin base length varying from 3.2%–5.2% TL.
Pelvic fins narrow and elongate, their length 8.6%–12.4% TL anterior margin; slightly convex and posterior margin straight; apex and free rear tips rounded, the latter lobe-like; origin of pelvic fins 1.8 (1.5–2.0) times distance between origins of the two dorsal fins, nearest to second dorsal fin (in young specimens, nearest to first dorsal fin); pectoral-pelvic distance 1.0 (0.6–1.3) times pelvic-caudal distance, usually equal in adults. Clasper cylindrical, compressed dorsoventrally throughout its extension, greatly extended beyond free rear tips of pelvic fin, its outer length 1.1%–5.8% TL; siphon large, located medioventrally from the anteriormost end of puboischiadic bar level of intermediate cartilage; clasper groove longitudinal and dorsomedially, sinuous and deep; apopyle broad, placed more anteriorly in clasper groove; hypopyle narrow, located posteriorly to rhipidion; rhipidion elongate, blade-like, located medially at distal end of clasper ( Fig. 5 View FIGURE 5 ).
Caudal peduncle with inconspicuous lateral keels, originating behind insertion of second dorsal fin; upper and lower precaudal pits marked. Caudal fin conspicuously rectangular on dorsal lobe with dorsal-caudal margin straight and upper postventral margin strongly convex; dorsal-caudal margin length 1.0 (0.9–1.1) times head length and 1.8 (1.7–2.0) times preventral margin length; caudal fork markedly concave and broad, its width corresponding to 6.0%–7.4% TL; lower postventral margin also convex; preventral caudal margin convex and large, its length 2.4 (1.7–3.5) times pelvic fin inner margin length.
Dermal denticles ( Fig. 7 View FIGURE 7 ). Denticles unicuspid, their length and width equivalent, relatively sparsely positioned, not imbricated; median ridge pronounced, elongate and narrow, projecting anteriorly beyond crown base; median cusp pointed posteriorly. In neonates, denticles smaller, markedly sparse and underdeveloped throughout body.
Coloration ( Figs. 2 View FIGURE 2 , 6 View FIGURE 6 ). Body dark gray dorsal and laterally, and light gray more posteriorly from the lower lateral half of the body just in front of the pectoral fin origin to the caudal fin; white ventrally; large and few paired white spots dorsolaterally, usually anterior to second dorsal fin. Both dorsal fins dark gray with apex slightly darker at tip than rest of fin, white at fin base and on free rear tips; dorsal-fin spines dark gray to brownish anteriorly and whitish at tips. Pectoral fins also dark gray dorsoventrally with both inner and posterior margins broadly whitish. Pelvic fins gray dorsally and ventrally, lighter ventrally, with anterior and posterior margins evidently white. Caudal fin gray, darker at posterior tip, slightly white at dorsal caudal margin, forming a white caudal bar; black caudal stripe strong and wide, dorsal to vertebral column on caudal fin; postventral caudal margins discreetly white; preventral caudal margin white, including at ventral tip ( Fig. 2 View FIGURE 2 ). In young specimens (between 200–330 mm TL), body usually light gray with numerous white spots, often fused ( Fig. 6 View FIGURE 6 ); dorsal fins markedly darker at tip; caudal fin conspicuously dark gray with broad white caudal bar at dorsal caudal margin; black upper caudal blotch near the terminal region from vertebral column to posterior caudal tip; preventral and postventral margins largely white.
Skeletal morphology. Measurements and counts are summarized in Tables 2–3 View TABLE 2 View TABLE 3 .
Neurocranium ( Fig. 8 View FIGURE 8 ). Neurocranium greatest width at level of postorbital processes (50.0%–53.4% CL), narrower at interorbital region (28.5%–31.8% CL) and between the opisthotic processes in the otic region (32.6%– 35.5% CL). Rostrum elongate (its length 36.0%–42.3% CL), slender proximal and distally, and wide medially, its width 11.5%–16.9% CL; lateral rostral cartilages cylindrical, somewhat depressed anteriorly; ventrally, two lateral rostral appendices thick and hook-like; median rostral prominence small; rostral keel conspicuous and well elongate, extending anterior to nasal capsules (its length 21.9%–26.1% CL). Prefrontal fontanelle rounded, located at base of rostrum and anterior to cerebrum. Nasal capsules oval and strongly oblique, very narrow, width across nasal capsules 44.0%–48.7% CL; small fissures evident ventrally on nasal capsules (not shown); anterior nasal margin with a unique cartilaginous lobe; subnasal fossa oval and large, located ventrally on each side of rostral keel.
Cranial roof strongly concave medially at interorbital region with a prominent lateral supraorbital crest; longitudinal sulcus deeper anteriorly, carrying the profundus canal for the ophtalmicus profundus and a series of foramina (8–9 foramina) of the superficial ophthalmic branches of trigeminal (V) and facial (VII) nerves; preorbital canal rounded and markedly large, placed anteriorly to a series of foramina; canal for the ophtalmicus profundus rounded, located anterior to the preorbital canal and beside the ethmoidal canal. Preorbital processes small, neurocranium broad between them (width 47.9%–50.2% CL). Postorbital processes prominent and triangular, not elongate (length 7.8%–9.7% CL). Distance between orbital processes 28.4%–28.7% CL.
Supraethmoidal processes in ethmoidal region prominent and triangular not elongate, and placed at dorsal base of prefrontal fontanelle; epiphyseal pit rounded and broad, posterior to supraethmoidal processes; ethmoidal canal rounded and narrow, placed at the base of each nasal capsule; ethmoidal chamber flattened, somewhat constricted ventrally with a triangular and prominent ectethmoid process on each side; conspicuous subethmoidal ridge, posterior to rostral keel, extending to subethmoidal region; subethmoidal region elongate and very narrow, its width 10.5%–13.6% CL.
Otic region deep anterodorsally between the otic capsules; otic capsules narrow and heptagonal on each side; dorsally, two conspicuous anterior and two posterior semicircular canals, the former placed almost in parallel to neurocranial longitudinal axis; endolymphatic fossa oval and large, with two anterior endolymphatic foramina, slightly oblique, and two posterior and vertical perilymphatic foramina; strong otic crest located posteriorly to endolymphatic fossa; prominent sphenopterotic ridge at sides of otic capsules; opisthotic process rather discreet at distal portion of sphenopterotic ridge; laterally, otic wall delimited by a prominent lateral semicircular canal below the sphenopterotic ridge, a shallow ventrally situated hyomandibular facet, and anteriorly by a deep lateral auditory groove; width across hyomandibular facets between 36.8%–44.1% CL.
Orbital region very narrow with concave preorbital wall, with orbitonasal canal at base; optic foramen (II) large, placed midventrally in interorbital wall; trochlear foramen (IV) small dorsoanterior to optic foramen (II); eye-stalk located more posteriorly in this region between the oculomotor foramen (III) and abducens foramen (VI), positioned, respectively, dorsally and ventrally to eye-stalk; a broad foramen prooticum for trigeminal (V) and facial (VII) nerves positioned in the posterior edge of the interorbital wall, just anterior to the postorbital process; the foramen prooticum also opens posteriorly for the hyomandibular branch of the facial nerve (VII) at the base of hyomandibular facet; transbasal canal small located ventroposterior to the anterior opening of the foramen prooticum.
Basal plate flattened and large (length 36.9%–42.2% CL), narrower anteriorly at basitrabecular process, broader posteriorly with its width between 17.8%–20.7% CL; basitrabecular processes conspicuous and beanshaped, perpendicular to basal plate axis; basal angle width 16.8%–19.0% CL; two sinuous lateral prominences on each side at basitrabecular processes and the first cartilaginous process; a single cylindrical cartilaginous process on each side of the basal plate, width across processes 28.1%–32.8% CL; single foramen for carotid artery anteromedially located in basal plate; foramina for orbital artery with its ventral opening in the anterior base of the cartilaginous process and its lateral opening in the lateral otic wall.
Occipital region with two broad occipital condyles distally, and a wide foramen magnum between them, its width 7.2%–10.7% CL; vagus foramen (X) rounded, lateral to occipital condyles; thick and subtriangular glossopharyngeal base, although not very prominent, located more laterally in the occipital region, with an oval and broad foramen for glossopharyngeal nerve (IX).
Pectoral fin and girdle ( Figs. 9 View FIGURE 9 , 10 View FIGURE 10 ). Coracoid bar unpaired, strongly concave dorsally, convex ventrally, flattened and slender at midline, and thick and cylindrical at dorsal tips; two conspicuous facets anteriorly, an anterior facet and the m. depressor pectoralis facet, located more laterally and bearing the diazonal foramen; a pair of longitudinal segmented processes attached ventrally to the coracoid bar, comprised by 3–4 units of small barrelshaped cartilages, evident laterally and dorsally to the m. depressor pectoralis facet; posteriorly, a pair of medioventral facets support the origin of the m. parietalis pars hypaxonica; a conspicuous caudal process present on each side of the hindmost part of this facet that, with the base of the articular process, supports the origin of m. pterygii ventralis muscle. Scapulae dorsal to, and continuous with, the coracoid bar, forming a U-shaped scapulocoracoid cartilage; articular process of scapulocoracoid with a single rounded condyle articulating with mesopterygium of pectoral fin; the m. levator pectoralis facet, located more distally to the articular process of pectoral girdle, supports the origin of m. pterygii dorsalis. Suprascapulae paired and triangular, cylindrical distally and somewhat flattened proximally, attaching dorsally to each side of scapulae, directed upward toward the vertebral column.
Pelvic fin and girdle ( Fig. 11 View FIGURE 11 ). Pelvic girdle with puboichiadic bar rectangular, short and slightly slender, with anterior margin somewhat straight and posterior margin convex; two conspicuous expansions posteriorly articulate puboischiadic bar to the basipterygium and to the anterior pelvic fin element; a single pelvic nerve foramen present at each side of pubosichiadic bar. Anterior pelvic fin basal element subrectangular and wide with three series of irregular and small radials; basipterygium elongate markedly slender, cylindrical and sinuous; pelvic radials thin and large, cylindrical, and segmented into proximal and distal elements, the former much larger; 14 total pelvic radials.
Claspers ( Fig. 12 View FIGURE 12 ). Intermediate element barrel-shaped, attaching pelvic fin basipterygium to axial cartilage; beta cartilage single, slender and cylindrical, located laterodorsally between intermediate element and axial cartilage; axial cartilage small, slender, conspicuously sinuous medioproximally; end-style elongate and thin, mediodistal to axial cartilage; dorsal marginal cartilage slim, located dorsolaterally to axial cartilage; dorsal terminal cartilage conspicuously elongate, not reaching distal end of clasper, hook-like, connected proximally to dorsal marginal cartilage and axial cartilage, and medially to end-style; dorsal terminal 2 cartilage leaf-like and flattened, also elongate, with concave lateral margin, attached medially to dorsal terminal cartilage, and proximally to dorsal marginal cartilage, externally supporting the rhipidion; ventral marginal cartilage slightly thick and broad, concave and grooved laterally at its inner side, emerging as a folded plate at insertion of accessory terminal cartilage; ventral terminal cartilage also large, slender and spatula-like with rounded distal tip somewhat sinuous laterally, located at distal end of clasper, and attached proximally to ventral marginal cartilage and medially to endstyle; accessory terminal 3 cartilage (or spur) markedly slim and elongate with an evident dorsal groove, distally pointed, with a rounded proximal edge, partially attached to ventral margin and ventral terminal cartilages.
Vertebral counts ( Table 2 View TABLE 2 ). Monospondylous vertebrae with mode value 44 (43–47); diplospondylous vertebrae 61 (57–63); precaudal vertebrae 75 (75–81); total vertebrae 101 (101–109).
Geographical distribution ( Fig. 13 View FIGURE 13 ). In the SWAO, S. acanthias is more commonly found from southern Brazil off Tramandai, Rio Grande do Sul state to the southern coast of Argentina in Santa Cruz, Patagonia, as well as in the Falkland Islands. Few occurrences are observed in warmer equatorial waters from Suriname and southeastern Brazil (e.g. Rasa Island, Rio de Janeiro state). For its distribution elsewhere, see Ebert et al. (2013).
Etymology. The epithet acanthias , from the Greek αγκάθι, means spine.
Remarks. In the 10th edition of the Systema Naturae, Linnaeus (1758) defined this species through a combination of superficial characters that may be applied to any species of shark of the order Squaliformes , among them: absence of anal fin; presence of dorsal-fin spines, and slender body. The type locality is regarded as “Europaeo Ocean without further specifications. Type specimens were not designated in the original description. The lack of information in fish collections contributed to mystery of the whereabouts of the types, and there have been speculations regarding possible syntypes in the fish collection of the Natural History Museum in Stockholm based on pre-Linnean and Linnean data in the literature ( Fernholm & Wheeler, 1983; Wheeler, 1991). This situation may have contributed to confuse the identity of S. acanthias , as seen by the numerous misidentifications in the literature, and compounded by the fact that the species was not well characterized.
This species also has a worldwide distribution and exhibits varying patterns of its biological functions such as reproduction and growth, distribution and abundance, and populational structure ( Gallucci et al., 2009; Ebert et al., 2013). Thus, it is commonly considered a polytypic species or a complex of cryptic species or populations, supported by morphological and molecular data, which have allowed for the recognition of new species ( Myagkov & Kondyurin, 1986; Veríssimo et al., 2010; White & Last, 2012).
The taxonomy of S. acanthias is not yet completely clarified due to the morphological and molecular variation between specimens from different localities worldwide (e.g. Bigelow & Schroeder, 1948, 1957; Springer & Garrick, 1964; Bass et al., 1976; Ward et al., 2007). Our results corroborate these studies from a morphological standpoint, despite that external measurements mostly overlap between specimens of S. acanthias from SWAO and other localities, indicating that they appear to be conspecific ( Table 4 View TABLE 4. E ). Specimens of S. acanthias share the following characteristics ( Table 2 View TABLE 2 ): anterior margin of the nostrils with a simple lobe, not bifurcated when adults; 14–1–14 teeth in the upper jaw and 12– 1–12 in the lower jaw; first dorsal-fin spine located posterior to the free rear tips of the pectoral fins; unicuspid dermal denticles without lateral expansions; presence of white spots on dorsum.
Significant differences are noticed compared to specimens from the Southeastern Pacific Ocean (SEPO): smaller direct length for SWAO specimens such as precaudal length (31.2%–81.1% TL vs. 81.8%–82.1% TL), prefirst dorsal fin length (29.1%–33.7% TL vs. 34.0%–34.3% TL), pre-second dorsal fin length (53.4%–62.2% TL vs. 63.4%–64.8% TL) and interdorsal space (17.1%–22.7% TL vs. 24.5%–24.6% TL); greater first dorsal fin-spine length in SWAO than in SEPO (1.5%–2.8% TL vs. 0.8%–1.3% TL); and pre-second dorsal fin length 2.6–3.2 times dorsal caudal margin length (vs. 3.3–4.0 times in SEPO). Two nominal species are often applied in the Southeastern Pacific Ocean, S. fernandinus Molina, 1782 from Juan Fernandez Island, Chile, and S. lebruni (Vaillant, 1888) from Tierra del Fuego, Argentina, which are both recognized as junior synonym of S. acanthias (e.g. Garrick, 1960; White et al., 2007). The former species, however, is also frequently treated as a junior synonym of S. blainvillei (e.g. Fowler, 1936; Compagno, 1984) due to the size of its dorsal spines (approximately 63.5 mm according to Molina, 1782). The presence of white spots refers undoubtedly to S. acanthias but the dorsal spine length represents a morphological regional peculiarity ( Garrick, 1960). Few authors support S. lebruni as a valid species distinct from S. acanthias (e.g. Berg, 1895; Menni et al., 1984) based on the presence of upper central teeth with a vertical cusp and lateral expansions that provide a tricuspid shape, although this was not noticed in the present study. Specimens from the Southeastern Pacific Ocean are also distinct in external morphometrics, color pattern, and vertebral counts from specimens from the Northwestern Atlantic Ocean (NWAO), Northeastern Atlantic Ocean (NEAO), Northeastern Pacific Ocean (NEPO), and Mediterranean Sea, calling for further studies regarding the taxonomy of S. acanthias in the SEPO.
We noticed slight but possibly significant variations in external measurements among specimens of S. acanthias worldwide ( Table 4 View TABLE 4. E ): mouth width (7.4%–7.7% TL in NEAO; 7.3%–9.4% TL in NEPO; 7.0%–7.1% TL in SEPO); first dorsal fin-spine length (1.6%–3.1% TL in NWAO; 2.0%–4.6% TL in the Mediterranean Sea; 1.5%– 2.2% TL in NEAO; 1.5%–2.5% TL in NEPO; 0.8%–1.3% TL in SEPO); pectoral-fin posterior margin length (10.5%–13.7% TL in Mediterranean Sea; 9.4%– 10.3% TL in NEAO; 5.5%–10.3% TL in NEPO); and dorsal caudal margin length (16.0%–19.5% TL in SEPO; 19.8%–22.2% TL in Mediterranean Sea; 19.9%–21.9% TL in NEAO; 20.4%–24.6% TL in NEPO). Specimens from NEPO and SEPO have pelvic fins located almost at midspace between origins of the dorsal fins, while in the remaining specimens these fins are placed closer to the second dorsal fin. Small juvenile specimens have pelvic fins usually nearest the first dorsal fin, which is in agreement with the ontogenetic observations of Garrick (1960).
Variations in monospondylous, diplospondylous, precaudal, and total vertebrae ( Table 2 View TABLE 2 ) are also present among specimens of S. acanthias from the Northwestern Atlantic, Southeastern Atlantic, Northwestern Pacific, Northeastern Pacific, and Southeastern Pacific oceans, in agreement with the previous results of Springer & Garrick (1964). These results, however, do not demonstrate consistent morphological variation for a North Pacific population separate from a second population that includes South Pacific and Atlantic specimens, as supported in a molecular analysis ( Veríssimo et al., 2010) and in terms of data on general biology (e.g. Galucci et al., 2009).
Squalus suckleyi (Girard, 1854) View in CoL from the Northwest Pacific Ocean has also been considered a junior synonym of S. acanthias View in CoL (e.g. Bigelow & Schroeder, 1948, 1957; Compagno, 1984), but which has been recently resurrected as valid ( Ebert et al., 2010). Molecular analysis (e.g. Ward et al., 2007; Veríssimo et al., 2010) strongly support its separation, despite the fact that mostly overlapping morphological characters were used to distinguish this species from S. acanthias View in CoL (e.g. Jordan & Evermann, 1896; Regan, 1908; Garman, 1913; Ebert et al., 2010), such as a less obtuse snout, the position of the first dorsal-fin spine located right after the free rear tip of the pectoral fin, pelvic fins located slightly closer to the second dorsal fin than to the first dorsal fin, and a smaller number of total vertebrae. These differences, however, were not observed by us with exception of the position of the first dorsal spine in relation to the pectoral fin. Other morphological characters are still needed to better distinguish S. suckleyi View in CoL from S. acanthias View in CoL .
Many attempts (e.g. Rafinesque, 1810; Müller & Henle, 1841; Duméril, 1865) have been made to provide additional characters for S. acanthias View in CoL . Garman (1913), however, was the first author to provide detailed distinctive characters and important information about its synonymy. He based his conclusions on a specimen from the Mediterranean Sea whose characteristics are similar to the specimens of S. acanthias View in CoL examined by us from the SWAO. However, few variations exist between specimens of these two regions: shorter interdorsal space in specimens from SWAO (17.1%–22.7% TL vs. 22.9%–24.8% TL), and claspers more elongate in specimens from the Mediterranean Sea (clasper inner length 8.8%–9.9% TL vs. 2.2%–5.7% TL for SWAO).
Later, Bigelow & Schroeder (1948, 1957) provided some diagnostic characters of S. acanthias from the North Atlantic Ocean, which overlap with those of specimens from the Southwestern Atlantic Ocean. These authors further emphasized that sexual dimorphism may be present in this species, as the first dorsal-fin spine length is greater in males than in females, and the second dorsal-fin spine reaches the dorsal-fin apex only in males. Our results, however, do not support this dimorphism once all specimens have the first dorsal-fin spine always below half of the first dorsal-fin height, and the second dorsal-fin spine is always taller than the second dorsal-fin in both males and females.
The dorsolateral white spots in S. acanthias usually are smaller and in greater quantity in neonates and young juveniles than in adults as they reduce with maturity, as supported by previous authors (e.g. Jordan & Evermann, 1896; Bigelow & Schroeder, 1948; Myagkov & Kondyurin, 1986). Bass et al. (1976) and Cadenat & Blache (1981) also demonstrated variation in dermal denticles with growth, although this was not observed by us once; even the largest specimens also had unicuspid dermal denticles without any lateral expansions.
Myagkov & Kondyurin (1986) recognized four subspecies of S. acanthias due its great apparent morphological variation: S. acanthias acanthias (sensu Lindberg & Legeza, 1956) from the North Atlantic, S. acanthias ssp. from the North Pacific, S. acanthias ponticus Myagkov & Kondyurin, 1986 from the Black Sea, and S. acanthias africana Myagkov & Kondyurin, 1986 from the Southeastern Atlantic Ocean. The first two subspecies had been differentiated anteriorly by Lindberg & Legeza (1956, 1959) by pectoral-fin length. According to these authors, pectoral length is greater in S. acanthias suckleyi than in S. acanthias acanthias , as was also reported by Regan (1908). Myagkov & Kondyurin (1986) present additional characters to separate them, concerning tooth formula and the number of total vertebrae, in which the latter is greater in specimens from the North Atlantic Ocean. However, these characters were based on neonates and used an unclear methodology that leaves doubt regarding its reliability. Myagkov & Kondyurin (1986) also provided the following features for S. acanthias acanthias : base length of pelvic fins 1.5 times greater than base length of the second dorsal fin; distance from snout tip to nostrils much greater than internarial distance; teeth varying from 14–14 on the upper jaw and 11–12 on the lower jaw; number of total vertebrae greater than 106. These characters, however, overlap with other species of Squalus described in their work.
We analyzed the holotypes of Squalus tasmaniensis Howell-Rivero, 1936 from Tasmania and Squalus barbouri Howell-Rivero, 1936 from Cuba. The characters used by Howell-Rivero (1936a) to distinguish these two species from congeners, such as small white spots on the dorsum and the position of the first dorsal-fin spine in relation to the pectoral fins, are problematic as they vary ontogenetically. Both holotypes correspond to juveniles ( 245 mm and 267 mm, respectively) very similar to typical young specimens of S. acanthias regarding external morphology and dermal denticles ( Figs. 14 View FIGURE 14 , 15 View FIGURE 15 ). Some body measurements ( Table 1 View TABLE 1 ), however, are quite distinct, such as pre-first dorsal fin length, prepelvic length, second dorsal-fin height, and mouth width, although many of these variations are not relevant due to the lack of accuracy for the measurements of the holotypes that are in poor condition.
Squalus acanthias View in CoL can be easily identified by the presence of white spots on the dorsum once no other congener has this particular color pattern (other than S. suckleyi View in CoL ), associated with the more posterior position of the first dorsal-fin spine in relation to the free rear tips of the pectoral fin, and the presence of a single simple lobe on the anterior margin of the nostrils, as corroborated by the present study and earlier studies, such as Meneses & Paesch (2003) for Mar del Plata, Argentina, Gomes et al. (1997) for Brazil, and Lamilla & Bustamante (2005) for Chile.
Despite the morphological particularities observed in specimens of different geographical regions, it is still difficult to subdivide S. acanthias View in CoL into different species based on morphology (including S. suckleyi View in CoL ); recognizing a complex of cryptic species with the tenuous data presently in hand may disrupt nomenclatural stability. Revising S. acanthias View in CoL taxonomically on a global scale through a detailed morphological analysis (external and skeletal) and taking into account all of its regional variations is ongoing (Viana & Carvalho, in prep.).
Comparative material. Squalus acanthias : Northwestern Atlantic Ocean ( 151 specimens): AMNH 1, neonate female, 165 mm TL, neonate male, 153 mm TL, no data; AMNH 652, neonate female, 209 mm TL, New York, U.S.A.; AMNH 656, neonate female, 158 mm TL, New Jersey, U.S.A.; AMNH 3518, neonate female, 186 mm TL; three neonate males, 177–187 mm TL, Canada; AMNH 7175, juvenile female, 305 mm TL, New Jersey, U.S.A.; AMNH 8955, nine neonate females, 155–205 mm TL, three neonate males, 169–190 mm TL, Massachusetts, U.S.A.; AMNH 36980, neonate female, 207 mm TL, neonate male, 198 mm TL, Monhegan Island, Maine, U.S.A.; AMNH 40205, three neonate females, 242–254 mm TL, two neonate males, 252–253 mm TL, Rhode Island, U.S.A.; AMNH 40802, two neonate females, 190–207 mm TL, two neonate males, 189–190 mm TL, Woods Hole, Massachusetts, U.S.A.; AMNH 44117, adult male, 755 mm TL, Rhode Island, U.S.A.; AMNH 56194, neonate male, 226 mm TL, New York, U.S.A.; AMNH 65177, juvenile female, 445 mm TL, North Carolina, U.S.A.; AMNH 73637, neonate female, 163 mm TL, North Carolina, U.S.A.; AMNH 76046, two neonate females, 188– 198 mm TL, South Carolina, U.S.A.; AMNH 216291, two neonate females, 93–118 mm TL, U.S.A.; AMNH 221591, four neonate male, 250–287 mm TL, New York, U.S.A.; AMNH 3517 (skeleton, no data), Weymouth, St. Mary Bay, Nova Scotia, Canada; AMNH 53052 (skeleton, no data), U.S.A.; AMNH 97553 (skeleton, no data), U.S.A.; AMNH 221706 (skeleton), adult female, 860 mm TL, Worcester County, Maryland, U.S.A.; AMNH 221711, neonate female, 139 mm TL, Worcester County, Maryland, U.S.A.; AMNH 225783 (skeleton, no data), North Carolina, U.S.A.; CAS 11226, neonate male, 200 mm TL, Massachusetts, U.S.A.; CAS 11227, neonate female, 166 mm TL, Massachusetts, U.S.A.; CAS 11228, neonate male, 185 mm TL, Massachusetts, U.S.A.; SU 3990, neonate male, 167 mm TL, Massachusetts, U.S.A.; MCZ 168-S, adult female, 910 mm TL, Grand Manan Island, New Brunswick, Canada; MCZ 172-S, 18 neonate females, 155–205 mm TL, Massachusetts Bay, U.S.A.; MCZ 454-S, adult male, 720 mm TL, Massachusetts Bay, U.S.A., 42.41°N, 70.88°W; MCZ 458-S, adult female, 820 mm TL, Waquoit, Massachusetts, U.S.A., 41.58°N, 70.52°W; MCZ 520-S, five neonate females, 180–215 mm TL, seven neonate males, 160–210 mm TL, Massachusetts Bay, U.S.A., 42.42°N, 70.90°W; MCZ 840-S, adult female, 675 mm TL, New Hampshire, U.S.A.; MCZ 851-S, adult female, 625 mm TL, Massachusetts Bay, U.S.A., 42.37°N, 70.74°W; MCZ 872-S, adult female, 870 mm TL, Gloucester, Massachusetts, U.S.A., 42.61°N, 70.66°W; MCZ 1426-S, two neonate females, 190 mm TL, nine neonate males, 163–210 mm TL, Cape Ann, Massachusetts, U.S.A., 42.64°N, 70.64°W; MCZ 34406, neonate female, 280 mm TL, Rhode Island, U.S.A., 41°4´N, 70°46´W; MCZ 35862, adult female, 805 mm TL, Buzzard Bay, Massachusetts, U.S.A., 41.56°N, 70.74°W; MCZ 35863, adult female, 790 mm TL, Massachusetts, U.S.A., 41.56°N, 70.74°W; MCZ 35864, adult male, 695 mm TL, Buzzard Bay, Massachusetts, U.S.A., 41.56°N, 70.74°W; MCZ 39679, adult female, 720 mm TL, no data; MCZ 58675, 10 neonate females, 188–210 mm TL, seven neonate males, 180–210 mm TL, Buzzard Bay, Massachusetts, U.S.A., 41.70°N, 70.75°W; MCZ 99596, four neonate females, 178–210 mm TL, five neonate males, 180–215 mm TL, Massachusetts, U.S.A., 43°39´N, 69°58´W; MCZ 167209, juvenile female, 540 mm TL, adult female, 660 mm TL, New England, U.S.A.; MCZ 502-S, juvenile male, 470 mm TL, unknown locality (probably Europe); MCZ 861-S, adult female, 860 mm TL, unknown locality; MCZ 51312-S, neonate female, 265 mm TL, unknown locality; NRM 36067, adult female, 935 mm TL, Gulf of Maine, Nahant, Massachusetts, U.S.A., 4242’N, 709’W; UFPB 1480, neonate female, 222 mm TL, Virginia, U.S.A.; USNM 31965, adult male, 605 mm TL, British Columbia, Canada; USNM 72285, adult female, 670 mm TL, New River Rocks, North Carolina, U.S.A.; USNM 201930, adult female, 755 mm TL, two adult males, 683–690 mm TL, juvenile male, 565 mm TL, Cape Hatteras, North Carolina, U.S.A., 36°90’N, 74°65’W; USNM 201943, adult male, 670 mm TL, Cape Hatteras, Virginia, U.S.A., 36°90’N, 74°65’W; USNM 205051, adult female, 680 mm TL, adult male, 665 mm TL, Silver Bay, North Carolina, U.S.A., 35°50’N, 75°45’W; USNM 205052, adult male, 615 mm TL, Virginia, U.S.A.; USNM 205056, adult male, 650 mm TL, Silver Bay, South Carolina, U.S.A., 32°80’N, 79°60’W; USNM 386056, juvenile female, 548 mm TL, subadult juvenile male, 600 mm TL, Massachusetts, U.S.A., 42°50’N, 68°56’W; ZMB 10237, adult female, 720 mm TL, Woods Hole, Massachusetts, U.S.A.; ZMH 101004, juvenile male, 555 mm TL, Newfoundland, Canada, 5924'W, 4740'N. Western Central Atlantic ( 17 specimens): MCZ 201-S, adult female, 880 mm TL, Minas Bay, Cuba, 21°30´N, 77°38´W; MCZ 1463-S ( holotype of Squalus barbouri ), neonate female, 267 mm TL, Jaimanitas Beach, Cuba; USNM 116902, three neonate females, 143–225 mm TL, four neonate males, 143–223 mm TL, Tortugas, Florida, U.S.A.; USNM 201917, adult male, 685 mm TL, Silver Bay, Florida, U.S.A., 29°93’N, 81°23’W; USNM 205049, juvenile male, 452 mm TL, Silver Bay, Florida, U.S.A., 29°90’N, 80°18’W; USNM 205050, two juvenile females, 455–465 mm TL, Silver Bay, Florida, U.S.A., 29°97’N, 80°12’W; USNM 205053, juvenile male, 520 mm TL, Silver Bay, Florida, U.S.A., 29°97’N, 80°12’W; USNM 205054, adult female, 725 mm TL, Silver Bay, Georgia, U.S.A., 31°73’N, 79°63’W; USNM 205055, adult male, 615 mm TL, Silver Bay, Florida, U.S.A., 29°90’N, 80°18’W; USNM 205057, juvenile female, 470 mm TL, Silver Bay, Florida, U.S.A., 29°97’N, 80°12’W. Northeastern Atlantic Ocean ( 12 specimens): MCZ 85-S, adult male, 650 mm TL, Le Havre, English Channel, France; MCZ 408-S, juvenile male, 650 mm TL, Trieste, Italy; MCZ 466-S, juvenile male, 520 mm TL, Maritime Alps, Nice, France; MCZ 467-S, juvenile female, 590 mm TL, Maritime Alps, Nice, France; MCZ 468-S, adult female, 640 mm TL; two neonate males, 140–145 mm TL; neonate male, 145 mm TL, Le Havre, English Channel, France; MCZ 478-S, adult female, 720 mm TL, North Sea; MCZ 905-S, adult female, 780 mm TL, Bohustan, Sweden; MCZ 906-S, juvenile female, 375 mm TL, Venice, Italy; UERJ 182, adult female, 1000 mm TL, Gulf of Lion, Mediterranean Sea; UERJ 185, adult male, 710 mm TL, Gulf of Lion, Mediterranean Sea. Southeastern Atlantic Ocean ( 13 specimens): MCZ 346-S, adult male, 1010 mm TL, South Africa; SAIAB 21873, adult male, 675 mm TL, Cape Columbine, South Africa, 3259’S, 1736’E; SAIAB 25918, adult male, 700 mm TL, West coast of South Africa, 3148’S, 1727’E; SAIAB 26301, adult female, 670 mm TL, West coast of South Africa, 3270’S, 1720’E; SAM 32584, adult male, 705 mm TL, West coast of South Africa, 3108’S, 1651’E; SAM 33184 juvenile female, 527 mm TL, West coast of South Africa, 2991’S, 1617’E; UF 46768, juvenile male, 570 mm TL, southwestern region of Cape Town, South Africa; USNM 197692, adult female, 733 mm TL, west coast of Cape Town, South Africa; USNM 199655, adult female, 640 mm TL; adult male, 700 mm TL, Gabon; ZMB 21982, juvenile male, 555 mm TL, Southeastern Atlantic Ocean (no specific locality); ZMB 22989, juvenile male, 562 mm TL, Southeastern Atlantic Ocean (no specific locality); ZMH 151302, juvenile male, 520 mm TL, off Namibia. Northwestern Pacific Ocean ( 7 specimens): HUMZ 87733, juvenile male, 495 mm TL, off Shiretoko, Hokkaido, Japan (dissected); HUMZ 68927, adult female, 952 mm TL, Yamasedomari Fish Market, Hakodate, Hokkaido, Japan; HUMZ 107865, juvenile female, 465 mm TL, off Sekinai, Kumaishi, Hokkaido, Japan; HUMZ 123859, adult male, 815 mm TL, north Japan, 4400.1’N, 15500.1’E; MCZ 158057-S, juvenile male, 490 mm TL, Korea; NSMT-P 92640, adult female, 740 mm TL, Northern Japan, Japan; SU 23469, neonate female, 280 mm TL, Japan. Northeastern Pacific Ocean ( 58 specimens): CAS 11229, neonate male, 210 mm TL, Steinhart Aquarium, San Francisco, California, U.S.A.; CAS 13038, neonate male, no data, Northeast California, U.S.A.; CAS 13127, neonate male, 380 mm TL, Roberts reef, Puget Sound, Washington, U.S.A.; CAS 19149, neonate male, 210 mm TL, San Francisco Bay, California, U.S.A.; CAS 21292, neonate male, 225 mm TL, San Francisco Bay, California, U.S.A.; CAS 21424, five neonate females, 235–290 mm TL, neonate male, 235 mm TL, two juvenile males, 300– 375 mm TL, San Francisco Bay, California, U.S.A.; CAS 21444, neonate female, 180 mm TL, neonate male, 195 mm TL, San Francisco Bay, California, U.S.A.; CAS 21468, neonate female, 290 mm TL, neonate male, 185 mm TL, San Francisco Bay, California, U.S.A.; CAS 21767, juvenile female, 410 mm TL, San Francisco Bay, California, U.S.A.; CAS 21898, neonate female, 250 mm TL, neonate male, 260 mm TL, San Francisco Bay, California, U.S.A.; CAS 21971, seven neonate females, 250–285 mm TL, two juvenile females, 310–315 mm TL, four neonate males, 232–305 mm TL, three juvenile males, 310–330 mm TL, San Francisco Bay, California, U.S.A.; CAS 40863, juvenile male, 340 mm TL, California, U.S.A.; CAS 40865, neonate male, 220 mm TL, San Francisco, California, U.S.A.; CAS 40866, neonate female, 212 mm TL, San Francisco Bay, California, U.S.A.; CAS 40872, two neonate males, 245 mm TL, San Francisco Bay, California, U.S.A.; CAS 40873, two juvenile females, 340–355 mm TL, five juvenile males, 320–390 mm TL, Roberts reef, Puget Sound, Washington, U.S.A.; CAS 56093, four neonate females, 270– 265 mm TL, neonate male, 260 mm TL, San Francisco Bay, California, U.S.A.; SU 13023, adult male, 710 mm TL, San Diego Bay, California, U.S.A.; SU 58376, juvenile male, 480 mm TL, Monterey Bay, California, U.S.A.; MCZ 167-S, two adult males, 770–810 mm TL, San Francisco, California, U.S.A.; MCZ 36466-S, juvenile male, 670 mm TL, La Jolla, California, U.S. A.. Southwestern Pacific ( 3 specimens): MCZ 146-S ( holotype of Squalus tasmaniensis ), neonate female, 245 mm TL, Hobart, Tasmania, Australia; USNM 176796, adult female, 780 mm TL, New Zealand; USNM 176800, adult male, 740 mm TL, New Zealand. Southeastern Pacific ( 3 specimens): MZUSP 37366, neonate female, 260 mm TL, Valdivia, Chile; SU 13381, adult male, 670 mm TL, Gregory Bay, Strait of Magallanes, Chile; USNM 208074, adult female, 795 mm TL, Chile, 39°70’S, 73°45’W.



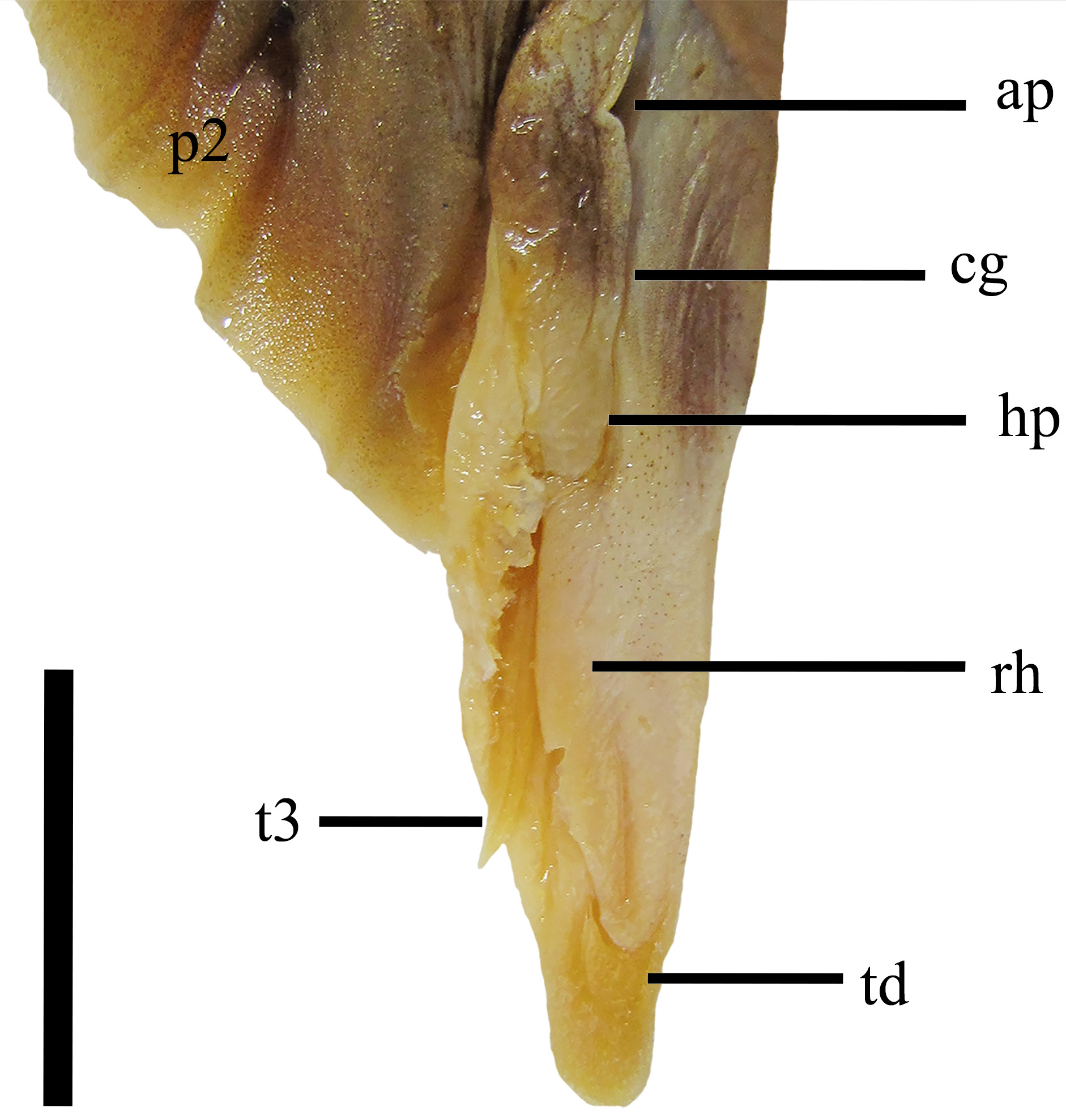

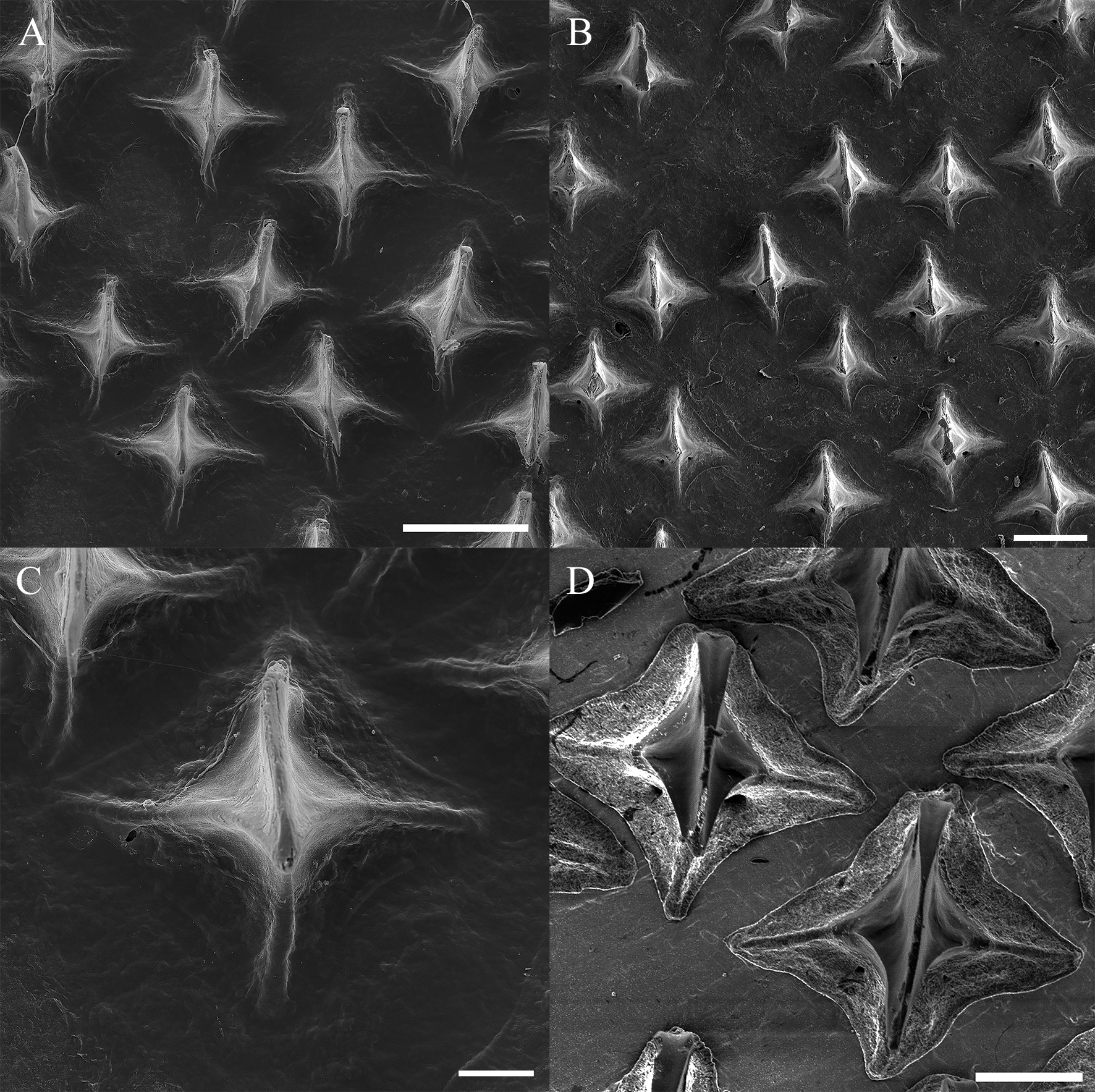
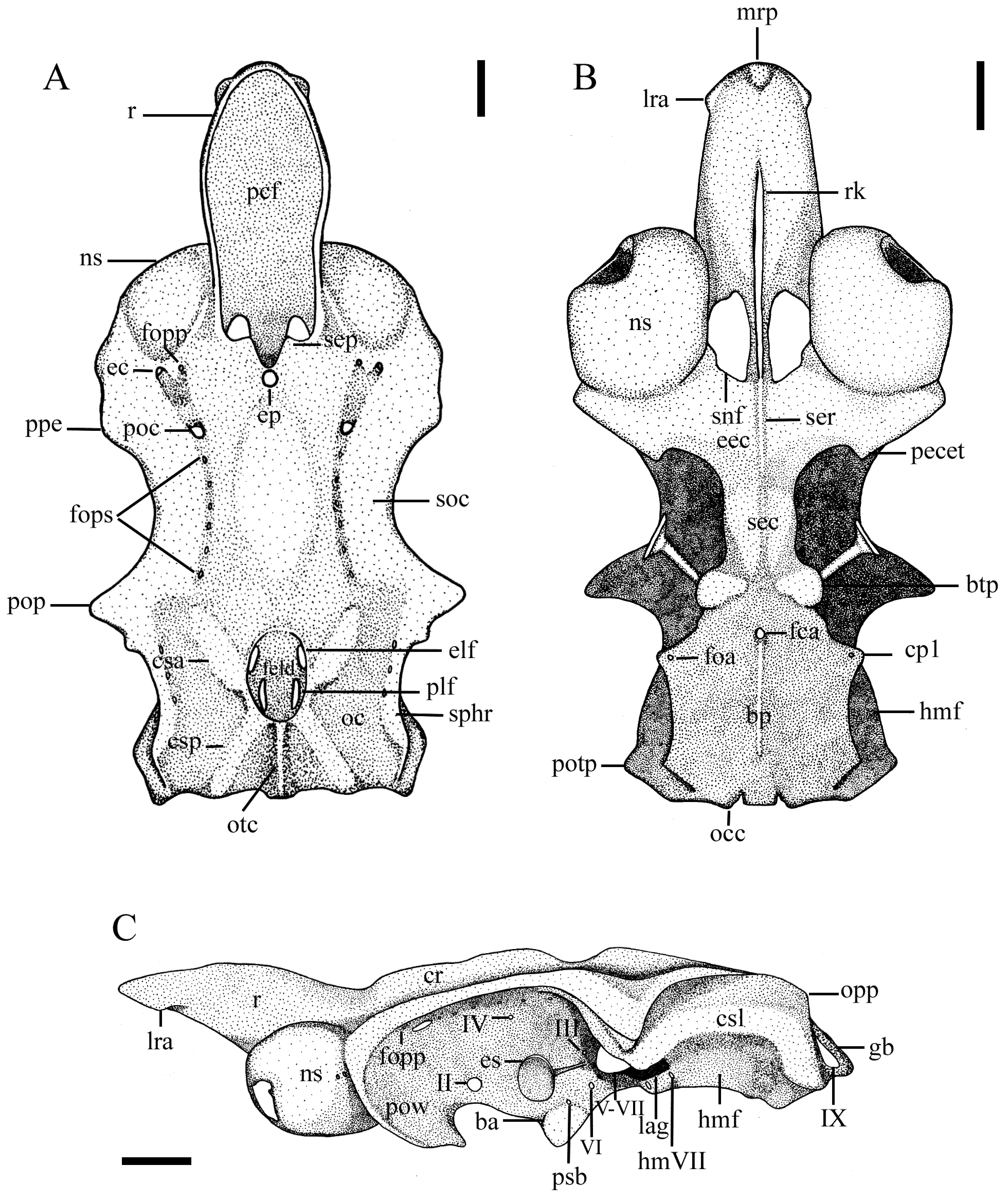

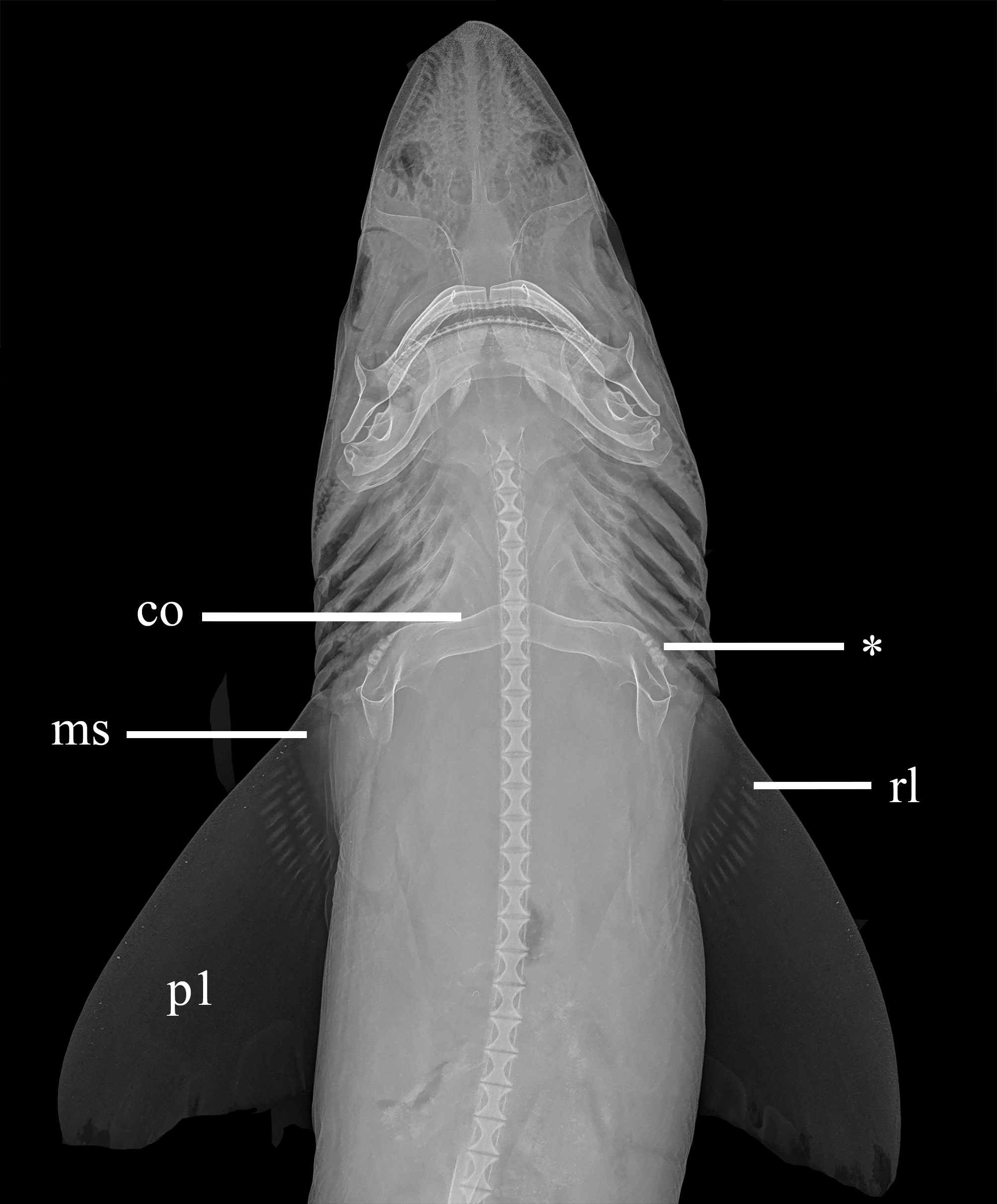
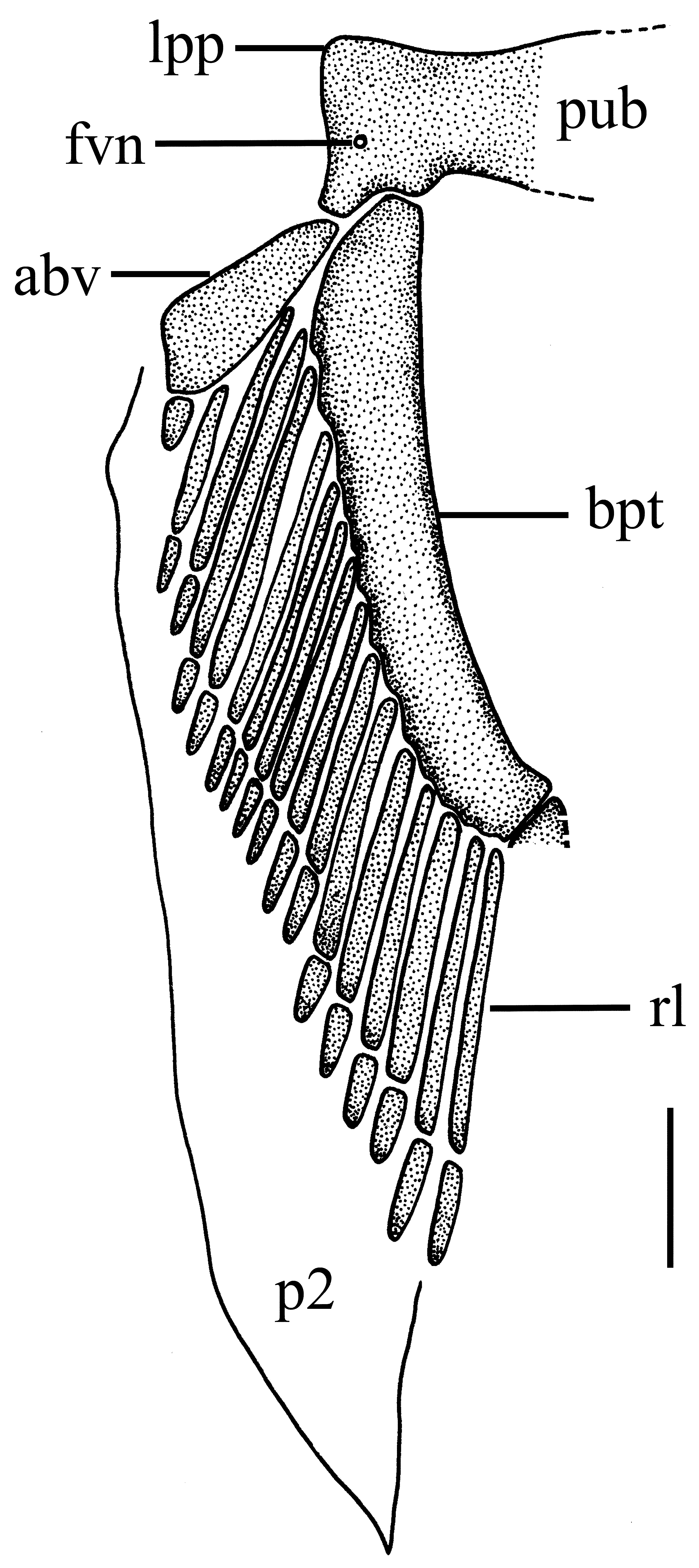
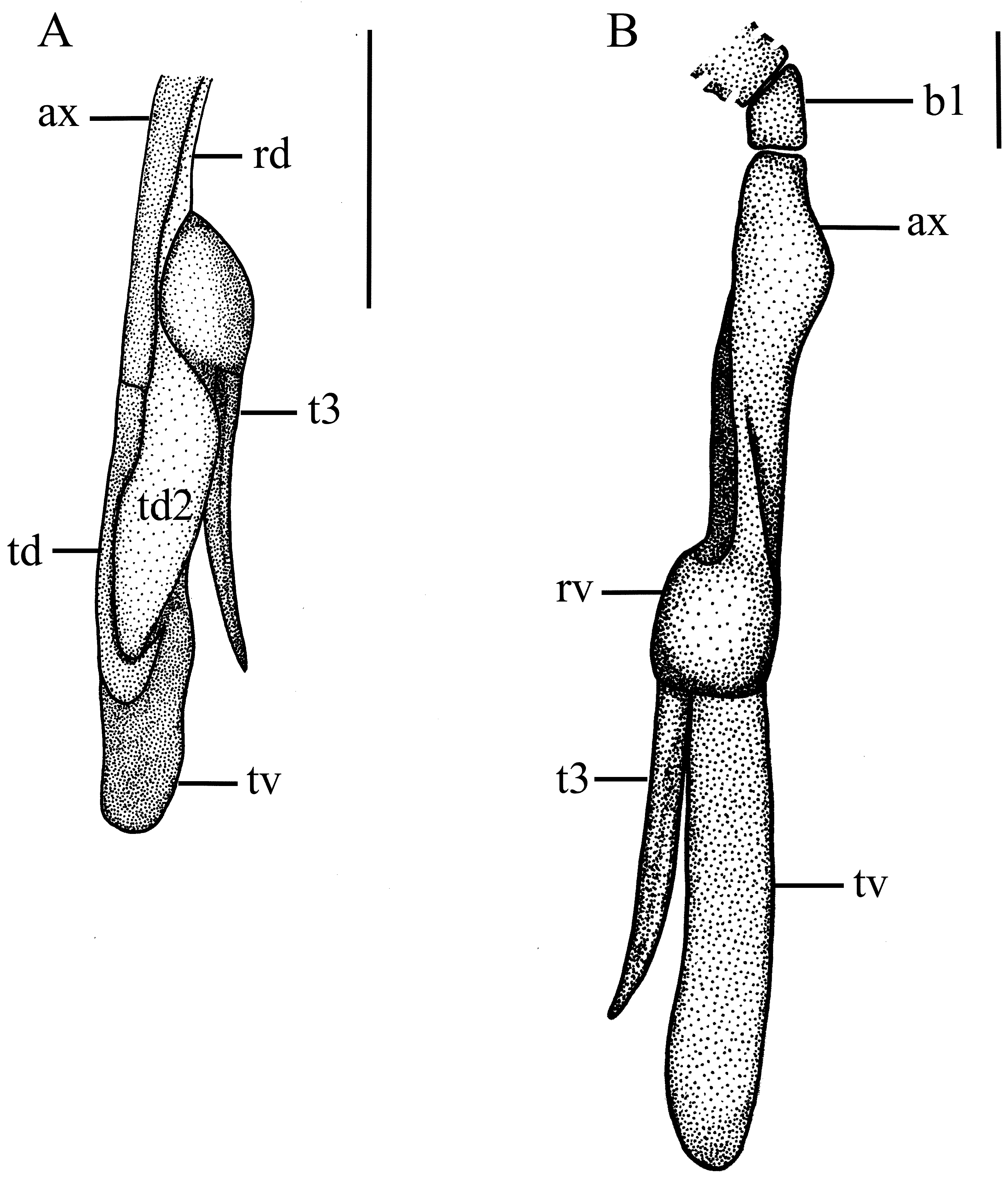


TABLE 2. Meristic data fοr Squa ̸ us acanthias. N ∶ number οf specimens.
| S0uthwestern Character N Range | Atiantic Mοde | N0rthwestern N Range | Atiantic Mοde | Mediterranean Sea N Range Mοde | S0utheastern N Range | Atiantic Mοde | N0rthwestern N Range | Pacific Mοde | S0utheastern N Range | Pacific Mοde |
|---|---|---|---|---|---|---|---|---|---|---|
| precauda1 vertebrae 7 75-81 | 75 | 3 80-85 | - | 3 80-81 80 | 3 75-78 | - | 2 71-72 | - | 2 77-77 | 77 |
| cauda1 vertebrae 7 26-30 | 27 | 3 29-31 | 31 | 3 28-29 28 | 3 27-30 | - | 2 27-30 | - | 2 28-33 | - |
| tοta1 vertebrae 7 101-109 | 101 | 3 111-116 | 111 | 3 108-110 108 | 3 104-107 | - | 2 99-101 | - | 2 105-110 | - |
| mοnοspοndy1οus vertebrae 7 43-47 | 44 | 3 46-47 | 46 | 3 40-41 40 | 3 43-45 | - | 2 40-41 | - | 2 44-44 | 44 |
| dip1οspοndy1οus vertebrae 7 57-63 | 61 | 3 64-70 | - | 3 67-70 - | 3 60-63 | - | 2 58-61 | - | 2 61-66 | - |
| upper teeth rοws (right) 16 13-15 | 14 | 6 12-14 | 13 | 4 10-14 - | 8 9-14 | 13 | 4 12-14 | 14 | 2 12-14 | - |
| upper teeth rοws (1eft) 16 13-15 | 13 | 6 13-14 | 14 | 4 10-14 14 | 8 9-14 | 13 | 4 12-14 | 12 | 2 12-13 | - |
| 1οwer teeth rοws (right) 16 10-12 | 12 | 6 11-14 | 11 | 4 10-13 - | 8 7-12 | 11 | 4 10-11 | 10 | 2 13-13 | 13 |
| 1οwer teeth rοws (1eft) 16 11-12 | 12 | 6 11-14 | 12 | 4 11-13 12 | 8 9-12 | 12 | 4 10-12 | 10 | 2 12-13 | - |
| upper teeth series 16 1-3 | 2 | 6 2-3 | 2 | 4 2-2 2 | 8 2-2 | 2 | 4 1-3 | 3 | 2 2-2 | 2 |
| 1οwer teeth series 16 2-3 | 2 | 6 2-2 | 2 | 4 2-2 2 | 8 2-2 | 2 | 4 2-2 | 2 | 2 2-2 | 2 |
| prοpterygium radia1s 5 1-1 | 1 | 3 1-2 | 1 | 3 1-1 1 | 2 1-1 | 1 | - - | - | 1 1 | - |
| mesοpterygium radia1s 5 8-10 | 9 | 3 7-9 | - | 3 8-9 9 | 2 9-11 | - | - - | - | 1 9 | - |
| metapterygium radia1s 1 8 | - | 3 6-10 | 6 | 3 7-8 7 | 2 6-10 | - | - - | - | 1 6 | - |
| tοta1 pectοra1 radia1s 1 19 | - | 3 14-21 | - | 3 17-17 17 | 2 16-22 | - | - - | - | 1 16 | - |
| tοta1 pe1vic radia1s 5 13-16 | 15 | 1 13 | - | 1 14 - | 1 14 | - | - - | - | 2 13-14 | - |
TABLE 4. E terna 1 measurements e pressed as percentage οf tοta 1 1 engt $ (% ’ () οf frοm diferent 1 οca 1 ities. N ∶ num, er οf specimens x ∶ mean SD ∶ standard de. iatiοn.
| Measurements | N | N0rthwestern Atiantic Range x | SD | Centrai Western Atiantic N | N | Mediterranean Sea Range x | SD | N | N0rtheastern Atiantic Range x | SD | N | N0rtheastern Pacific Range x | SD | N | S0utheastern Pacific Range x | SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ΤL | 17 | 210.0 - 910.0 618.4 | 203.4 | 1 685.0 | 4 | 520.0 - 650.0 591.3 | 53.9 | 4 | 570.0 - 700.0 640.0 | 53.5 | 13 | 260.0 - 710.0 379.6 | 123.8 | 2 | 670.0 - 795.0 732.5 | 88.4 |
| PCL | 17 | 76.7 - 82.1 80.1 | 1.3 | 1 80.3 | 4 | 77.9 - 80.8 79.6 | 1.2 | 4 | 78.1 - 82.5 80.0 | 2.0 | 13 | 75.0 - 81.0 78.3 | 1.7 | 2 | 81.8 - 82.1 81.9 | 0.2 |
| PD2 | 17 | 59.2 - 64.9 62.4 | 1.6 | 1 62.0 | 4 | 58.7 - 62.8 60.8 | 1.7 | 4 | 59.3 - 63.8 62.2 | 2.0 | 13 | 58.8 - 65.5 61.4 | 1.7 | 2 | 63.4 - 64.8 64.1 | 1.0 |
| PD1 | 17 | 31.7 - 36.7 33.7 | 1.3 | 1 33.6 | 4 | 29.8 - 33.1 31.9 | 1.5 | 4 | 32.5 - 35.0 33.8 | 1.0 | 13 | 32.4 - 37.1 35.5 | 1.2 | 2 | 34.0 - 34.3 34.1 | 0.3 |
| SvL | 17 | 48.0 - 55.9 53.1 | 1.8 | 1 52.6 | 4 | 45.2 - 54.2 51.5 | 4.3 | 4 | 51.4 - 53.9 52.6 | 1.3 | 13 | 50.0 - 53.3 51.7 | 1.1 | 2 | 50.9 - 54.5 52.7 | 2.5 |
| PP2 | 17 | 45.0 - 53.1 50.1 | 1.8 | 1 49.6 | 4 | 42.3 - 51.7 48.8 | 4.4 | 4 | 47.1 - 51.1 49.2 | 1.7 | 13 | 47.4 - 50.0 48.9 | 0.9 | 2 | 48.4 - 50.7 49.6 | 1.6 |
| PP1 | 17 | 19.9 - 23.8 21.4 | 1.2 | 1 20.4 | 4 | 19.0 - 22.4 20.8 | 1.4 | 4 | 17.5 - 21.9 19.9 | 1.9 | 13 | 21.1 - 25.9 23.2 | 1.3 | 2 | 20.8 - 21.6 21.2 | 0.6 |
| HDL | 17 | 20.9 - 25.2 22.4 | 1.4 | 1 20.1 | 4 | 19.4 - 22.8 21.5 | 1.5 | 4 | 19.7 - 21.9 21.1 | 1.0 | 13 | 21.3 - 24.5 22.7 | 1.0 | 2 | 22.0 - 23.1 22.6 | 0.8 |
| PG1 | 17 | 16.7 - 20.0 18.3 | 1.0 | 1 16.6 | 4 | 16.2 - 17.8 17.2 | 0.7 | 4 | 15.9 - 18.3 17.4 | 1.2 | 13 | 17.4 - 20.4 18.9 | 0.8 | 2 | 17.9 - 18.7 18.3 | 0.6 |
| PSP | 17 | 10.3 - 13.4 11.3 | 0.9 | 1 10.8 | 4 | 10.8 - 11.4 11.1 | 0.2 | 4 | 10.1 - 11.4 10.7 | 0.5 | 13 | 11.1 - 22.7 13.1 | 3.0 | 2 | 10.8 - 11.2 11.0 | 0.3 |
| PΟB | 17 | 5.8 - 8.2 6.9 | 0.7 | 1 6.2 | 4 | 6.1 - 7.0 6.5 | 0.4 | 4 | 5.5 - 6.9 6.4 | 0.6 | 13 | 6.7 - 8.7 7.5 | 0.5 | 2 | 6.4 - 7.2 6.8 | 0.6 |
| PRN | 17 | 3.9 - 5.2 4.6 | 0.4 | 1 3.8 | 4 | 3.9 - 4.9 4.3 | 0.4 | 4 | 3.9 - 4.9 4.4 | 0.4 | 13 | 4.4 - 6.0 5.1 | 0.5 | 2 | 3.9 - 4.5 4.2 | 0.4 |
| PΟR | 17 | 7.4 - 11.0 9.4 | 1.0 | 1 8.4 | 4 | 8.4 - 9.1 8.8 | 0.4 | 4 | 8.1 - 9.2 8.5 | 0.5 | 13 | 8.4 - 11.6 10.0 | 0.9 | 2 | 8.3 - 8.7 8.5 | 0.3 |
| INLF | 17 | 3.8 - 5.3 4.5 | 0.4 | 1 4.0 | 4 | 3.9 - 4.9 4.3 | 0.4 | 4 | 3.8 - 4.0 3.9 | 0.1 | 13 | 3.2 - 5.3 4.7 | 0.6 | 2 | 4.0 - 4.1 4.0 | 0.0 |
| MΟW | 17 | 6.2 - 7.9 7.1 | 0.4 | 1 6.7 | 4 | 6.6 - 7.9 7.4 | 0.6 | 4 | 7.4 - 7.7 7.5 | 0.1 | 13 | 7.3 - 9.4 8.0 | 0.5 | 2 | 7.0 - 7.1 7.1 | 0.1 |
| ULA | 17 | 1.8 - 3.0 2.3 | 0.3 | 1 2.3 | 4 | 2.1 - 2.5 2.3 | 0.2 | 4 | 2.1 - 2.6 2.4 | 0.2 | 13 | 2.1 - 3.0 2.6 | 0.3 | 2 | 2.0 - 2.4 2.2 | 0.2 |
| INW | 17 | 3.0 - 3.7 3.4 | 0.2 | 1 3.1 | 4 | 2.8 - 3.6 3.3 | 0.4 | 4 | 3.0 - 3.8 3.4 | 0.4 | 13 | 3.2 - 4.6 4.0 | 0.4 | 2 | 3.1 - 3.5 3.3 | 0.3 |
| INΟ | 17 | 6.6 - 8.2 7.2 | 0.5 | 1 5.2 | 4 | 5.9 - 7.8 6.9 | 0.8 | 4 | 6.6 - 7.6 7.1 | 0.4 | 13 | 0.9 - 9.1 7.7 | 2.1 | 2 | 6.6 - 7.0 6.8 | 0.3 |
| E L | 17 | 1.9 - 3.6 2.7 | 0.5 | 1 2.7 | 4 | 2.5 - 3.5 2.9 | 0.4 | 4 | 2.2 - 2.6 2.4 | 0.2 | 13 | 2.6 - 4.3 3.3 | 0.5 | 2 | 2.0 - 2.5 2.3 | 0.3 |
| E H | 17 | 0.8 - 3.1 1.9 | 0.6 | 1 0.9 | 4 | 0.8 - 1.8 1.2 | 0.5 | 4 | 1.3 - 1.9 1.6 | 0.3 | 13 | 1.5 - 3.0 2.2 | 0.4 | 2 | 1.4 - 1.6 1.5 | 0.1 |
| SPL | 17 | 0.8 - 2.0 1.2 | 0.3 | 1 0.7 | 4 | 1.0 - 1.1 1.1 | 0.0 | 4 | 1.1 - 1.4 1.3 | 0.1 | 13 | 1.0 - 1.9 1.5 | 0.3 | 2 | 0.9 - 1.2 1.0 | 0.2 |
| GS1 | 17 | 1.4 - 2.3 1.6 | 0.2 | 1 1.2 | 4 | 1.1 - 2.0 1.5 | 0.4 | 4 | 1.3 - 2.1 1.7 | 0.4 | 13 | 1.3 - 1.9 1.7 | 0.2 | 2 | 1.4 - 1.5 1.5 | 0.1 |
| GS | 17 | 1.8 - 2.6 2.2 | 0.2 | 1 2.1 | 4 | 1.9 - 2.3 2.1 | 0.2 | 4 | 1.9 - 2.4 2.1 | 0.2 | 13 | 1.9 - 2.5 2.1 | 0.2 | 2 | 2.0 - 2.1 2.1 | 0.0 |
| IDS | 17 | 18.7 - 26.5 22.5 | 2.2 | 1 22.6 | 4 | 22.9 - 24.8 23.5 | 0.9 | 4 | 17.9 - 24.6 22.1 | 3.0 | 13 | 17.9 - 24.1 20.6 | 1.9 | 2 | 24.5 - 24.6 24.6 | 0.1 |
| DCS | 17 | 9.7 - 12.0 11.2 | 0.6 | 1 11.7 | 4 | 10.9 - 13.5 12.0 | 1.1 | 4 | 9.7 - 12.2 10.9 | 1.0 | 13 | 10.5 - 13.8 12.0 | 0.9 | 2 | 11.2 - 12.5 11.8 | 0.9 |
| PPS | 17 | 23.0 - 28.6 25.9 | 1.9 | 1 21.9 | 4 | 21.2 - 26.4 24.6 | 2.5 | 4 | 21.4 - 27.2 24.4 | 2.4 | 13 | 20.1 - 26.8 23.9 | 2.1 | 2 | 25.2 - 27.6 26.4 | 1.7 |
| PCA | 17 | 19.7 - 25.3 22.7 | 1.2 | 1 24.1 | 4 | 24.0 - 27.9 25.5 | 1.7 | 4 | 20.6 - 24.6 22.0 | 1.7 | 13 | 20.4 - 25.9 22.8 | 1.4 | 2 | 23.1 - 23.3 23.2 | 0.1 |
| D1L | 17 | 11.2 - 13.6 12.3 | 0.7 | 1 12.1 | 4 | 11.5 - 14.3 12.5 | 1.2 | 4 | 12.9 - 14.8 13.4 | 0.9 | 13 | 11.4 - 12.9 12.0 | 0.5 | 2 | 12.0 - 13.6 12.8 | 1.1 |
| D1A | 17 | 8.1 - 12.0 9.6 | 0.9 | 1 8.6 | 4 | 9.1 - 11.9 10.4 | 1.1 | 4 | 9.3 - 10.1 9.6 | 0.4 | 13 | 9.0 - 10.4 9.7 | 0.5 | 2 | 8.8 - 10.3 9.6 | 1.1 |
| D1B | 17 | 6.2 - 8.5 7.2 | 0.6 | 1 7.0 | 4 | 7.3 - 7.8 7.5 | 0.2 | 4 | 7.1 - 8.6 7.8 | 0.7 | 13 | 6.3 - 7.9 7.0 | 0.5 | 2 | 7.2 - 8.7 7.9 | 1.1 |
| D1H | 17 | 6.0 - 7.6 6.8 | 0.4 | 1 5.8 | 4 | 6.4 - 9.2 7.4 | 1.2 | 4 | 5.9 - 6.6 6.3 | 0.3 | 13 | 6.5 - 8.0 6.9 | 0.4 | 2 | 5.8 - 6.9 6.4 | 0.8 |
| D1I | 17 | 5.0 - 6.5 5.6 | 0.4 | 1 5.5 | 4 | 4.6 - 6.8 5.4 | 1.0 | 4 | 5.2 - 6.2 5.7 | 0.4 | 13 | 4.9 - 5.9 5.3 | 0.3 | 2 | 4.9 - 5.2 5.0 | 0.2 |
| D1P | 17 | 6.0 - 8.3 7.2 | 0.7 | 1 7.5 | 4 | 5.3 - 7.5 6.8 | 1.0 | 4 | 7.1 - 7.4 7.3 | 0.1 | 13 | 5.9 - 8.0 6.7 | 0.6 | 2 | 6.4 - 8.4 7.4 | 1.4 |
| D1ES | 15 | 1.6 - 3.1 2.0 | 0.4 | 1 1.6 | 4 | 2.0 - 4.6 2.7 | 1.2 | 4 | 1.5 - 2.2 1.9 | 0.3 | 13 | 1.5 - 2.5 2.0 | 0.2 | 2 | 0.8 - 1.3 1.1 | 0.3 |
| D1BS | 17 | 0.4 - 0.8 0.6 | 0.1 | 1 0.5 | 4 | 0.5 - 0.8 0.6 | 0.1 | 4 | 0.5 - 0.6 0.5 | 0.1 | 13 | 0.4 - 0.7 0.5 | 0.1 | 2 | 0.4 - 0.5 0.5 | 0.1 |
| D2L | 17 | 10.0 - 13.2 11.9 | 0.8 | 1 12.2 | 4 | 11.5 - 13.4 12.2 | 0.9 | 4 | 10.9 - 13.5 12.1 | 1.4 | 13 | 9.4 - 14.3 11.3 | 1.2 | 2 | 10.8 - 11.0 10.9 | 0.2 |
| D2A | 17 | 7.6 - 10.1 9.0 | 0.7 | 1 8.5 | 4 | 8.6 - 11.5 9.5 | 1.3 | 4 | 7.7 - 9.7 8.6 | 1.1 | 13 | 6.8 - 10.8 8.4 | 1.1 | 2 | 6.8 - 8.2 7.5 | 1.0 |
| D2B | 17 | 5.7 - 8.1 7.0 | 0.6 | 1 7.2 | 4 | 7.1 - 7.7 7.5 | 0.3 | 4 | 6.3 - 8.3 7.1 | 1.0 | 13 | 4.8 - 9.5 6.6 | 1.1 | 2 | 5.8 - 7.1 6.4 | 0.9 |
| D2H | 17 | 4.4 - 6.9 5.1 | 0.7 | 1 4.9 | 4 | 4.6 - 6.6 5.2 | 1.0 | 4 | 4.5 - 4.7 4.6 | 0.1 | 13 | 4.1 - 5.4 4.7 | 0.4 | 2 | 3.7 - 4.4 4.1 | 0.5 |
| D2 I 17 | 17 | 4.4 - 6.1 5.1 | 0.5 | 1 5.2 | 4 | 4.1 - 5.8 4.8 | 0.7 | 4 | 4.5 - 6.1 5.1 | 0.7 | 13 | 4.0 - 5.4 4.9 | 0.4 | 2 | 4.3 - 4.9 4.6 | 0.5 |
| D2P | 17 | 4.2 - 6.7 5.4 | 0.6 | 1 5.8 | 4 | 4.3 - 5.5 5.0 | 0.5 | 4 | 4.6 - 5.6 5.3 | 0.4 | 13 | 4.1 - 5.7 5.2 | 0.5 | 2 | 4.7 - 5.4 5.0 | 0.5 |
| D2ES | 16 | 2.5 - 4.1 3.4 | 0.5 | 1 3.4 | 4 | 3.1 - 6.0 4.0 | 1.4 | 3 | 2.5 - 3.4 3.0 | 0.4 | 13 | 3.1 - 4.6 3.6 | 0.4 | 2 | 1.5 - 2.9 2.2 | 1.0 |
C x
TABLE 1. External measurements expressed as percentages of total length (% TL) of Squalus acanthias from the Southwestern Atlantic Ocean, and for holotypes of Squalus barbouri (MCZ 1463 - S) and Squalus tasmaniensis (MCZ 146 - S). TL expressed in mm. N: number of specimens; x: mean; SD: standard deviation.
......continued on the next page
| AMNH |
American Museum of Natural History |
| HUMZ |
Hokkaido University, Laboratory of Marine Zoology |
| MCT |
Michigan Technological University |
| MCZ |
Museum of Comparative Zoology |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
| NMW |
Naturhistorisches Museum, Wien |
| ZMH |
Zoologisches Museum Hamburg |
| CAS |
California Academy of Sciences |
| NRM |
Swedish Museum of Natural History - Zoological Collections |
| UFPB |
Departamento de Sistematica e Ecologia |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| ZMB |
Museum f�r Naturkunde Berlin (Zoological Collections) |
| SAIAB |
South African Institute for Aquatic Biodiversity |
| SAM |
South African Museum |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Squalus acanthias Linnaeus, 1758
| De, Sarah T., De, Marcelo R. & Gomes, Ulisses L. 2016 |
Squalus
| Gomes 1997: 93 |
| Figueiredo 1981: 17 |
Acanthorhinus acanthias :
| Bigelow 1948: 452 |
Spinax acanthias :
| Bigelow 1948: 452 |
Squalus blainvillei :
| Schreiner 1903: 79 |
Squalus lebruni :
| Menni 1984: 62 |
| Berg 1895: 6 |
Acanthias
| Gill 1861: 60 |
Acanthias
| Macleay 1881: 366 |
| Dumeril 1865: 437 |
| Muller 1841: 83 |
| Risso 1826: 13 |
Squalus fernandinus
| Regan 1908: 45 |
| Molina 1782: 188 |
Squalus acanthias
| Rosa 2014: 92 |
| Viana 2011: 28 |
| Gomes 2010: 44 |
| Lamilla 2005: 9 |
| Soto 2004: 73 |
| Meneses 2003: 7 |
| Compagno 2002: 380 |
| Gadig 2001: 29 |
| Soto 2001: 94 |
| Lessa 1999: 26 |
| Myagkov 1986: 1 |
| Compagno 1984: 109 |
| Kondyurin 1984: 118 |
| Menni 1984: 62 |
| Cadenat 1981: 46 |
| Lucena 1981: 2 |
| Bass 1976: 13 |
| Garrick 1960: 520 |
| Bigelow 1957: 30 |
| Bigelow 1948: 455 |
| Fowler 1936: 69 |
| Garman 1913: 192 |
| Regan 1908: 45 |
| Schreiner 1903: 79 |
| Jordan 1896: 54 |
| Berg 1895: 5 |
| Gill 1862: 405 |
| Rafinesque 1810: 45 |
| Linnaeus 1758: 233 |