PACHYOSTEOMORPHI STENSIÖ, 1944
(Suborder
Brachythoraci Gross 1932
, in part; suborder
Pachyosteina Stensiö 1944
; suborder
Pachyosteomorphi Stensiö 1944
, in part;
Pachyosteomorphi Stensiö 1959
; order
Pachyosteida Obruchev 1964
; suborder
Pachyostei Stensiö 1969
)
The
Pachyosteomorphi
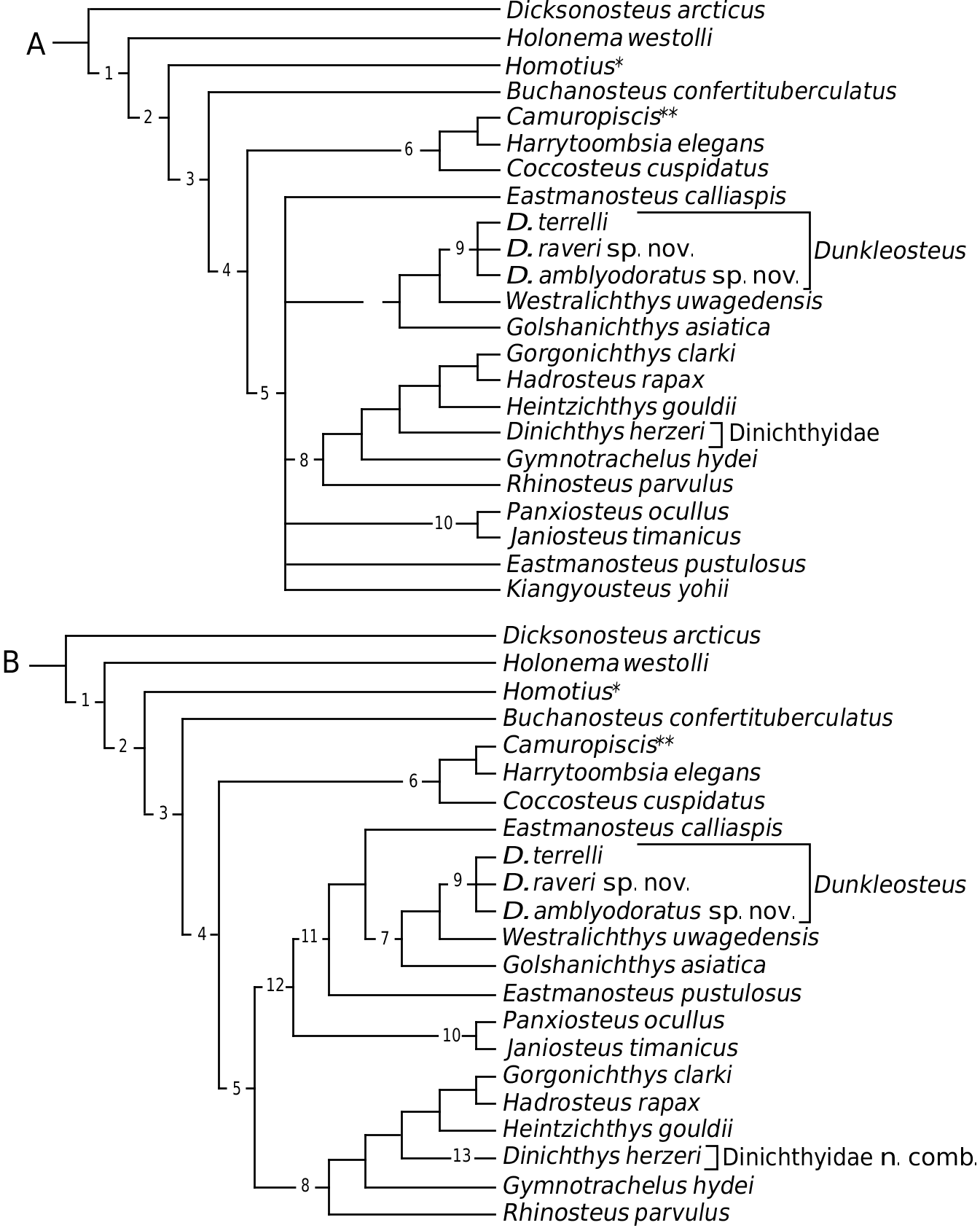
(node 5; Fig. 9
View Figure 9
) are a subclade within the Eubrachythoraci (node 4; Fig. 9
View Figure 9
), and are sister group to the Coccosteomorphi (node 6; Fig. 9
View Figure 9
). The following families are included here within the Eubrachythoraci
Pachyosteomorphi
: Braunosteidae Stensiö, 1959 (included within the
Selenosteidae sensu Denison, 1978
),
Bungartiidae Denison, 1975
,
Dinichthyidae
,
Dunkleosteidae
,
Hadrosteidae Gross, 1932
,
Leiosteidae Stensiö, 1963
,
Mylostomatidae Woodward, 1891
,
Panxiosteidae Wang, 1979
,
Selenosteidae
(including the
Pachyosteidae Gross, 1932
and the
Rhinosteidae Stensiö, 1963
),
Titanichthyidae Dean, 1901b
,
Trematosteidae Gross, 1932
,
Brachydeiridae Gross, 1932
,
Leptosteidae Jaekel, 1911
, and
Rachiosteidae Stensiö, 1963
(the inclusion of the latter three families is tentatively based on their opened pectoral fenestra in some members of each family, and the probable loss of the spinal plate); and Coccosteomorphi –
Camuropiscidae
,
Coccosteidae Traquair, 1888
,
Incisoscutidae Denison, 1984
, Torosteidae Gardiner & Miles, 1990 (with the latter included within the
Plourdosteidae
by Gardiner & Miles, 1994; Long 1995).
The various definitions of the
Pachyosteomorphi
result in the confusion of group membership. Stensiö (1944) first defined the
Pachyosteomorphi
as all the non-coccosteomorph
Brachythoraci
. He did not provide a clear diagnosis for the group, but did include some forms like
Di. herzeri
, the Pachy- osteidae, and the
Trematosteidae
. Stensiö (1959) later included the
Pachyosteomorphi
in the Aspinothoracidi, based on the pectoral fin and endoskeletal girdle structure alone. To account for the presence of a dermal plate connecting the lateral and ventral thoracic armour in
Dunkleosteus, Stensiö
coined the term ‘pseudospinal’ for what he thought was an independently derived spinal plate. As pointed out by Miles & Dennis (1979), Stensiö assumed the absence of spinals to be primitive. It then appears clear that, based on a parsimony argument, the ‘pseudospinal’ of Stensiö represents a homologous and primitive feature of the
Pachyosteomorphi
( Mark-Kurik, 1963; Heintz, 1968; Carr, 1991). The reduction of the spinal plate associated with the loss of a spinal pit is a synapomorphy for the
Pachyosteomorphi
( Gardiner & Miles, 1990: their character 22.32, although the latter character is equivocal because of missing data; in the present analysis, character 46). Unambiguous synapomorphies for the Eubrachythoraci in the present study include: (1) the development of a continuous occipital thickening (character 12, state 1); (2) the subequal length of the preorbital plate relative to the central plate (character 15, state 0); (3) a reduction in the ossification of the palatoquadrate forming two ossifications (autopalatine and quadrate; character 57, state 1); (4) the loss of the groove for the postorbital sensory line on the suborbital plate (character 81, state 0); and (5) the confluence of the postorbital branch of the infraorbital sensory line and supraoral sensory line grooves (character 83, state 1). The
Pachyosteomorphi
are characterized by: (1) the increased length of the central–nuchal plate contact (character 21, state 1); (2) the presence of a short and broad median dorsal plate (character 36, state 0); (3) the development of a lateral contact between the suborbital and preorbital plates (character 54, state 1); (4) the presence of posterolateral processes on the parasphenoid (character 72, state 1); and (5) the loss of the groove for the ventrolateral sensory line on the anterior lateral plate (character 77, state 0).