
Polypterid
|
publication ID |
https://doi.org/10.26028/cybium/2020-441-003 |
|
DOI |
https://doi.org/10.5281/zenodo.10881473 |
|
persistent identifier |
https://treatment.plazi.org/id/304B075D-6865-E85F-FCB5-8730FD1CFAEE |
|
treatment provided by |
Felipe |
|
scientific name |
Polypterid |
| status |
|
Comparative morphology of the finlet spines in Polypterid species ( Figs 13-18 View Figure 13 View Figure 14 View Figure 15 View Figure 16 View Figure 17 View Figure 18 )
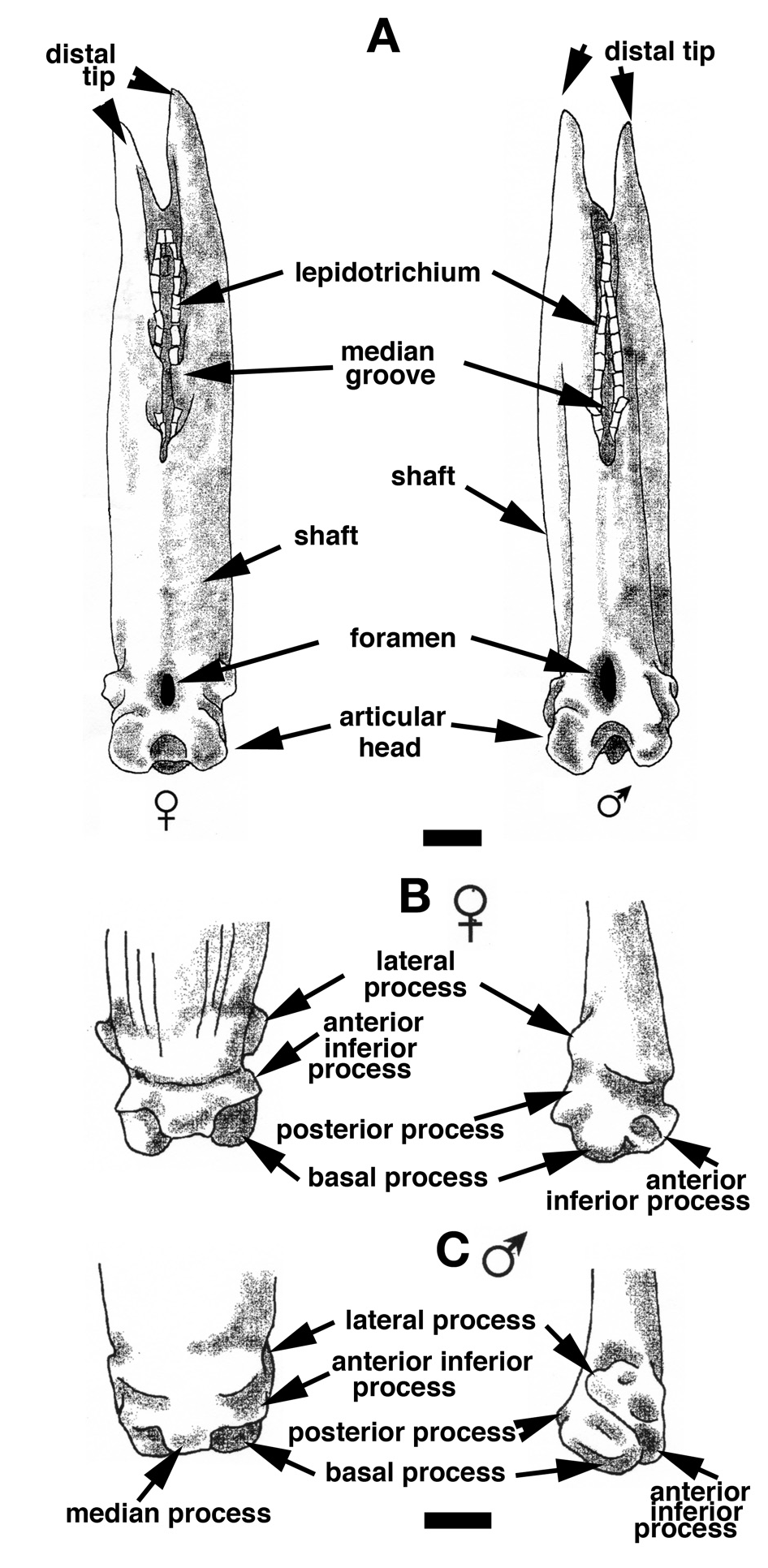
Comparisons of the first finlet spine have been done for all the species listed; the whole set of spines has been compared for a portion of the species (one to five fish according the species availability). In general, their global organization is similar to that described for Polypterus senegalus . However, there are several significant differences, in particularly, ones concerning the respective development and the relative position of the processes on the articular head of the spine.
For the four species of which males and females have been compared ( P. delhezi , P. ornatipinnis , P. senegalus and E. calabaricus ), we observed a common sexual dimorphism in the shape of the spine shaft: narrow with lateral depressions for the male and wide for the female. The age and sex of the specimen drastically influence the appearance of the ganoine layer. Indeed, the dentine is more widespread for larger specimens, and negatively correlated to the ganoine development. Thus, caution is required when using the anterior face of the spine for species identification. In juveniles, the ganoine is limited to a thin pad on the edges of the finlet spine and it progressively extends to irregular patches on the anterior face. At the first spine in young adults, the ganoine layer may cover the entire anterior surface to the upper transversal pad. The adult P. senegalus , P. delhezi , P. palmas , P. ornatipinnis and P. polli show this juvenile characteristic, but the ganoine slightly regresses on the posterior spines to the median line where the finlet web attaches. At the first spine of an adult P. retropinnis , the ganoine layer covers the whole surface except for the basal median region (similar to the last spine of P. senegalus ). In P. bichir adults, the ganoine is reduced from the first spine to ridges separated by the dentine. Finally, for adult P. ansorgei and P. endlicheri , the ganoine covers only a very small surface ( Fig. 13 View Figure 13 ).
Most species show strait shafts similar to those of Polypterus senegalus , except in P. ornatipinnis and P. retropinnis , for which the shaft is slightly curved, and in P. bichir katangae , P. b. lapradei and P. weeksii , for which it is strongly curved. From the lateral view, figure 14 shows that there is minute morphological variation of the articular head of spines. Only P. retropinnis stands out because its lateral process is clearly more spread out ( Fig. 14 View Figure 14 ). Therefore, it is not a particularly powerful character for discriminating species.
In posterior view, the region surrounding the lepidotrichium branches insertion is quite different among the Polypterus species ( Fig. 15 View Figure 15 ). For most of them, like in P. senegalus , the groove is continuous and the bony planes that integrate the branches are almost never fused. When they are fused, they form a pore, which disappears soon after basal bridge development. The opening of the groove is either regular without any constrictions as for the female P. senegalus ( Figs 11A View Figure 11 , 15 View Figure 15 ) or irregular with several constrictions as for the male of P. bichir ( Fig. 15 View Figure 15 ). For P. ansorgei and for P. ornatipinnis , the groove is always divided into two unequal parts separated by a bony bridge. For the three subspecies of P. bichir , each bridge closes and forms several depressions from which the three first branches of the lepidotrichium set out. However, the groove does not split between the last secondary branch and the main branch.
At first glance, the number of lepidotrichium has no limited factor other than the height of the spine. But for the adult fishes, this number seems to reach a limit between three and five; we observed one specimen of Polypterus senegalus ( 274 mm TL) with six branches. The formation of the branches, as drawn by Daget (1950: fig. 50), is erroneous – the median case does not exist; for all the finlets the spine is already completely formed with its unique lepidotrichium. The secondary branches form later from the primary lepidotrichium. However, we do confirm Sewertzoff (1924) and Daget (1950) observations that the secondary branches are never bifurcated.
The square outline of the articular head of the most anterior finlet has been observed for all species except Polypterus ansorgei , P. bichir bichir , and P. endlicheri ( Fig. 16 View Figure 16 ). Indeed, for P. ansorgei , a deletion of the lateral processes and a gap between the basal processes give the articular head a triangular shaped outline ( Fig. 16 View Figure 16 ). In P. b. bichir and P. endlicheri , the articular head is trapezoidal because the lateral processes are less developed ( Fig. 16 View Figure 16 ). Interestingly, even though the articular head of P. polli is square, it differs in that the posterior and the basal processes are fused and the lateral processes are slightly developed; then its articular head appears soft and rounded, which resembles the rounded articular head of Erpetoichthys ( Fig. 16 View Figure 16 ). In spite of the morphological variation noted among the whole set of finlet spines, we observed that in all Polypterus (except for P. ansorgei due to a lack of samples) the articular head shape is preserved.
In Polypterus retropinnis , the lateral processes are located in a more posterior position, far outside the posterior margin of the shaft ( Fig. 16 View Figure 16 ). The lower-posterior margin of the articular head may be round or concave in its median region depending on whether the basal and posterior processes have merged or not. A round and clean margin (almost straight) is often seen for the first spines of most of the adult species except P. ansorgei , P. bichir katangae and to a certain extent for P. b. lapradei, P. b. bichir and P. ornatipinnis ( Fig. 16 View Figure 16 ). In P. b. bichir , the first spine shows a straight margin, whereas the ultimate finlet spine is strongly concave. However, for the other species, the developmental stage of the spine does not seem influential.
The basal surface of the articular head of finlet spines can also be registered in a typical geometrical shape ( Fig. 17 View Figure 17 ): i) rectangular ( P. ansorgei , P. bichir , P. delhezi (not illustrated), P. retropinnis ; ii) square: P. ornatipinnis , P. palmas , P. weeksi ; iii) round: P. polli and E. calabaricus . These various shapes are the result of the differential development of the processes.
The histological comparative study of the spiny ray ground cross-sections does not show any specific histological characteristics ( Fig. 18 View Figure 18 ). They are all built on the model of Polypterus senegalus ( Meunier, 1980) . There are minor differences, which seem to be more correlated to the height of the spine than to a species diagnosis. The lower part of the spine is organized around the medullar cavity, from which the vascular canals set out and form an extended network. The spine is bordered at its lateral and posterior margin by dentine tissue and/or bone tissue, depending on whether it is close to the articular head or not. While we move away from the base of the spine, the medullar cavity disappears where the branches of the lepidotrichium set out. There is no longer any bony tissue, except for the bone from the inserted hemi-segments of the lepidotrichium; the composition of the spine shaft is reduced to dentine and ganoine ( Fig. 18 View Figure 18 ) ( Meunier, 1980; Meinke, 1982). The anterior area is composed of dentine with ganoine patches. These patches are in fact fused odontodes with few dispersed bony elements clustered around osteocytes. Thus, this part of the spine can be considered to be composed of osteodentin ( Ørvig, 1967). As one moves to the surface of the spine, the ganoine patches are more and more developed and closer to each other; they can even fuse, especially in larger individuals ( Fig. 18 View Figure 18 ), and form the so-called odonto-complex ( Ørvig, 1977).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Order |
|
|
Family |