
Anadia mcdiarmidi, Kok & Rivas, 2011
|
publication ID |
https://doi.org/ 10.5852/ejt.2011.3 |
|
DOI |
https://doi.org/10.5281/zenodo.3717557 |
|
persistent identifier |
https://treatment.plazi.org/id/327B87E6-6162-FFCB-FDB4-447DFC5CFCEB |
|
treatment provided by |
Plazi |
|
scientific name |
Anadia mcdiarmidi |
| status |
sp. nov. |
Anadia mcdiarmidi sp. nov.
Figs 1-7 View Fig View Fig View Fig View Fig View Fig View Fig , Table 1
Anadia breweri (nomen nudum) – Gorzula 1992: 276, figs 168-171
Anadia species a – Gorzula & Señaris 1999: 114-115, fig. 88
Anadia sp. A – McDiarmid & Donnelly 2005: 514
Anadia breweri (nomen nudum) – Brewer-Carías & Audy 2010: 210, unnumbered figure
Holotype
IRSNB 2677 View Materials (field number PK 3565 ), 1 adult ♂ collected by Philippe J. R. Kok, 03-05-2011 at 11h39, summit of Abakapá-tepui , Bolívar State, Venezuela, 05°11'09''N 062°17'36''W, 2201 m elevation. GoogleMaps
Paratopotypes
(n=3) 1 adult ♂ ( IRSNB 2674 , field number PK 3578 ) collected by Philippe J.R. Kok, 06-05-2011 at 9h40, summit of Abakapá-tepui , Bolívar State, Venezuela, 05°10'50''N 062°17'49''W, 2242 m elevation); GoogleMaps 1 juvenile ♂ ( IRSNB 2675 , field number PK 3577 ) collected by Philippe J.R. Kok, 06-05-2011 at 9h10, summit of Abakapá-tepui , Bolívar State, Venezuela, 05°11'00''N 062°17'34''W, 2200 m elevation GoogleMaps ; and 1 other juvenile ♂ ( IRSNB 2676 , field number PK 3589 ) collected by Philippe J.R. Kok, 09-05-2011 at 9h00, summit of Abakapá-tepui , Bolívar State, Venezuela, 05°11'07''N 062°17'19''W, 2204 m elevation. GoogleMaps
Etymology
The specific epithet is a noun in the genitive case, honouring Roy McDiarmid, North American herpetologist (Smithsonian Institution), for his contribution to the knowledge of the Pantepui herpetofauna.
Generic allocation
The new species is placed in the genus Anadia because it fits the 11 diagnostic traits for the genus proposed by Oftedal (1974): dorsal scales smooth, essentially homogeneous; no distinct rows of much smaller scales between lateral scales; ventral scales smooth, quadrangular; head scales smooth, without striation; single frontonasal separating nasals; paired prefrontals; several large pigmented palpebrals; gular scales smooth, essentially homogeneous, with no distinctly enlarged longitudinal rows; 7-10 large preanal scales [maximum 9 in Oftedal (1974)]; femoral pores conspicuous in adult males; and limbs well developed, pentadactyl with all toes clawed.
Definition and diagnosis
A fairly robust species currently not assigned to any species group. The new species is characterized by the following combination of characters: (1) size small, body robust (maximum known SVL 62.2 mm); (2) tail longer than SVL; (3) dorsal scales small, quadrangular; (4) middorsal scales 53-57; (5) suboculars large, unequal in size, with sometimes one scale slightly protruding downward between 4 th and 5 th supralabial, suboculars separated from palpebrals by 2-3 rows of mostly inconspicuous tiny scales, a few of them distinctly enlarged with angular downward protrusion between suboculars; (6) nasal entire, without sub-nostril groove; (7) 3-4 large pigmented palpebrals; (8) body uniform beige or greyish to bluish brown in life, devoid of any conspicuous pattern in males; (9) absence of any dark line on side of head in adult males; (10) venter immaculate golden grey in life; (11) femoral pores 9-10 on each side in males, unknown in females; (12) preanal pores absent; (13) hemipenis globose, weakly bilobed, bordered by numerous flounces (>20, including about eight medioproximal asulcate flounces)
bearing comblike rows of minute weakly mineralized spinules, sulcus spermaticus divided in the crotch by a small fleshy bumplike structure, sulcus branches barely detectable.
The new taxon is immediately distinguished from Anadia ocellata , A. vittata , A. rhombifera , and A. petersi [all members of the ocellata group of Oftedal (1974)] and A. bogotensis [single member of the bogotensis group of Oftedal (1974)] in having quadrangular dorsal scales (subhexagonal in species of the ocellata group, imbricate in A. bogotensis ), and in lacking a subnostril groove or divided nasal (nasal divided or subnostril groove present in species of the ocellata group and in A. bogotensis ); from A. altaserrania [a member of the bitaeniata group of Oftedal (1974) according to Harris & Ayala (1987)] in having prefrontals (lacking in A. altaserrania ), in having more femoral pores in males (9-10 in A. mcdiarmidi vs. 3-4 in A. altaserrania ), and in having a higher number of middorsal scales (53-57 in A. mcdiarmidi vs. 45-47 in A. altaserrania ); from A. bitaeniata , A. brevifrontalis , A. hobarti , A. pamplonensis , and A. pulchella [all members of the bitaeniata group of Oftedal (1974) according to La Marca & García-Pérez (1990)] in having a higher number of middorsal scales (53-57 in A. mcdiarmidi vs. maximum 50 in species of the bitaeniata group); from A. blakei and A. marmorata [members of the marmorata group of Oftedal (1974)] in having a higher number of middorsal scales (more than 50 in A. mcdiarmidi vs. less than 35 in A. blakei and A. marmorata ); from A. bumanguesa [a member of the bitaeniata group of Oftedal (1974) according to Rueda-Almonacid & Caicedo (2004), but a possible synonym of A. steyeri and thus a member of the steyeri group of Oftedal (1974) according to Rivas et al. (in press)] in having a lower number of scales around midbody (35-36 in A. mcdiarmidi vs. 41 in A. steyeri ), and in having pigmented palpebrals (unpigmented in A. bumanguesa ); from A. pariaensis (not assigned to any species group in the original description) in having a much lower number of middorsal scales (54- 57 in A. mcdiarmidi vs. 70-72 in A. pariaensis ); from A. steyeri [single member of the steyeri group of Oftedal (1974)] in having a lower number of scales around midbody (35-36 in A. mcdiarmidi vs. 40-45 in A. steyeri ); and from A. escalerae (not assigned to any species group in the original description), the geographically closest relative, in having a robust body (slender in A. escalerae ), in having smooth dorsal scales (weakly keeled in A. escalerae ), in lacking dorsolateral stripes (present in A. escalerae ), in having a lower number of middorsal scales (54-57 in A. mcdiarmidi vs. 63 in A. escalerae ), and in having a lower number of scales around midbody (35-36 in A. mcdiarmidi vs. 40 in A. escalerae ).
Description of the Holotype
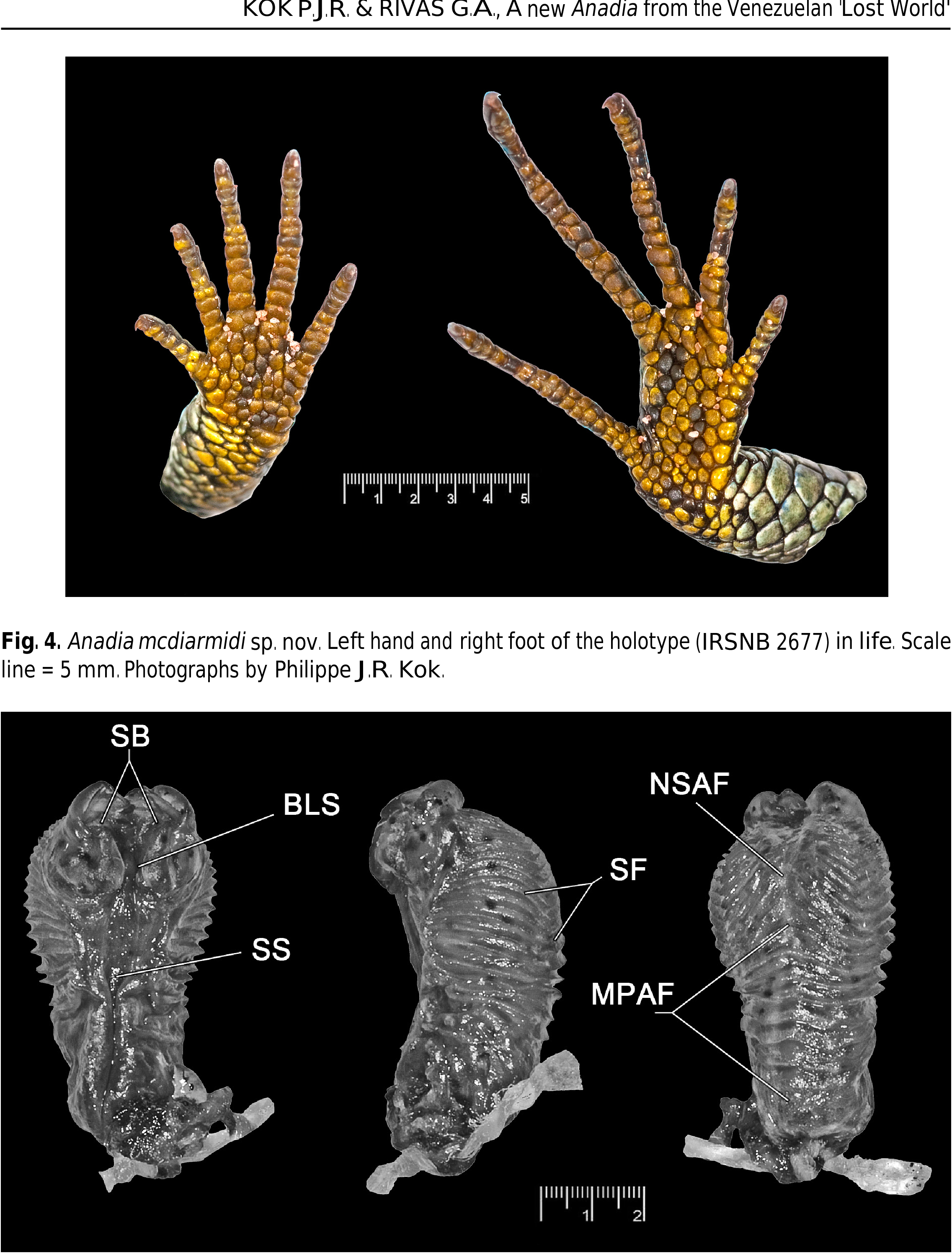
An adult male in good condition ( Figs 1A, C View Fig , 2 View Fig , 3 View Fig , 4, 6), with 62.2 mm SVL and 110.0 mm TL (see also Table 1). A rather robust Anadia species with moderately long limbs (digits overlap when limbs are adpressed along body). Snout attenuate, flat in profile. HL 25% of SVL, 1.5 times longer than wide, 1.6 times wider than high; head conspicuously wider than neck. Neck long, 68% of HL, 35% of AXG. SAL 93% of AXG, 46% of SVL. Body wider than deep. Tail dorsoventrally flattened, about 1.8 times longer than SVL. Limbs pentadactyl with all digits clawed. Forelimb 27% of SVL, 52% of AXG; hind limb 36% of SVL, 73% of AXG.
Scutellation on the dorsal surface of head is typical for the genus [see fig. 1 in Oftedal (1974)], except the condition of the parietals and interparietal (see below and Figs 2 View Fig , 7 View Fig ). The frontal is damaged, the frontoparietals are slightly damaged. Head scales are smooth with a few minute scattered pits, those pits are especially concentrated on the upper and posterior margins of supralabials, on the lower and posterior margins of infralabials, and on the posterior margin of rostral.
Rostral much wider than deep, laterally in contact with nasals and first supralabial, dorsally in contact with large frontonasal. Frontonasal with nearly straight anterior margin, posteriorly pointed. Paired prefrontals in contact, medial suture short. Frontal damaged, but obviously wider anteriorly and with blunt anterior point. Paired frontoparietals slightly damaged, with long medial suture, in contact with interparietal, parietals, and two posterior supraoculars. Three large supraoculars subequal in size. Interparietal divided into about two scales unequal in size. Parietals not extending as far as interparietal posteriorly, divided into two scales on the left side, into four scales on the right side. A series of six small to large occipitals (postparietals) circling the common posterior margin of parietals and interparietal.
Nasal scale entire, with a small indentation situated anterodorsally to nostril on the left side. Nasal scale in contact with rostral, posteriorly in contact with prefrontal, and in broad contact with loreal. Loreal large, pentagonal, in broad contact with first superciliary (= presuperciliary) and frenocular, in point contact with preocular. One small preocular, three postoculars. First superciliary large, followed by four shorter superciliaries. No small azygous scale between superciliaries and supraoculars. Frenocular followed posteriorly by three suboculars. Third subocular slightly extending to lip between supralabials 4-5 on the left side only. Suboculars separated from palpebrals by 2-3 poorly defined rows of mostly inconspicuous tiny scales, two of them distinctly enlarged with angular downward protrusion between suboculars. Seven supralabials.
Eight ciliaries along upper eyelid. Lower eyelid scales translucent, slightly pigmented, with four higherthan-wide palpebrals.
Temporal scales subimbricate, smooth, with flat surfaces, slightly larger above and smaller below. Ear opening broad, ovoid, slightly inclined posterodorsally, edged with small, slightly pebblelike scales; tympanum recessed, pigmented.
Underside of head with six infralabials on each side. A large mental followed by a large postmental in contact with first two infralabials. Two pairs of large genials in broad contact medially, in lateral contact with infralabials 3-4; two pair of postgenials, the first pair large, each scale in contact with last genial and infralabials 4-5, the second pair much smaller, each scale in contact with first postgenial and in point contact with fifth infralabial. Gulars arranged in transverse rows, becoming slightly larger posteriad, culminating in a well-defined collar row of eight subequal scales; no distinctly enlarged longitudinal rows. Side of neck between ear and collar pebbled with subequal irregularly shaped juxtaposed scales.
Middorsal scales 54. Dorsal scales on neck subimbricate, irregularly shaped, with rounded surfaces. Dorsal body scales smooth, juxtaposed, mainly quadrangular, a few irregularly shaped, but most are longer than wide, in transverse rows only. Lateral scales smooth, similar to dorsals, but smaller.
Ventral scales much wider than dorsals, smooth, juxtaposed, quadrangular, slightly longer than wide except for ca. four median longitudinal rows of square to wider than long scales on belly; in 14 longitudinal rows at midbody and 31 transverse rows between collar and vent.
Eight marginal preanal scales anterior to vent, the middle two narrower than the adjacent lateral ones, most outer ones very small. Femoral pores well developed, in swollen pore scales mostly in linear contact; 9 pores on left thigh, 10 on right. Pores do not extend onto preanal area ( Fig. 3 View Fig ).
Caudal scales smooth, subimbricate, rectangular, in transverse rows around tail.
Scales on dorsal surfaces of arm large, smooth, imbricate; ventral surfaces of arm with similar, but smaller and less imbricate scales. Hind limbs with large, smooth subimbricate to imbricate scales on anterior face of thighs and on lower legs; scales similar but smaller on ventral side of thigh. Dorsal and posterior sides of thigh with much smaller, irregularly shaped juxtaposed scales.
Moderate-size, imbricate scales atop hands and feet. Supradigital scales single; upper and lower ungualsheath scales covering base of claws, leaving tips well exposed. Palms and soles with small slightly raised juxtaposed scales (Fig. 4). Two enlarged smooth thenar scales at base of pollex. Subdigital lamellae mostly single. Lamellae under first (I) through fifth (V) finger (right/left side): I: 7/6, II: 9/9, III: 11/11, IV: 13/14, V: 10/9. Lamellae under first (I) through fifth (V) toe (right/left side): I: 6/6, II: 10/10, III: 14/12, IV: 17/15, V: 15/6 (large part of Toe V missing on left side).
Tongue lanceolate; proximal two-third unpigmented, distal one-third heavily pigmented, including fork. Upper surface behind fork covered with imbricate scalelike papillae that extend onto ventrolateral sides of tongue. Raised medioventral side of tongue with numerous (>10) of thin, oblique, anteriorly converging and pointed (chevronlike) infralingual plicae; anterior pair of plicae swollen, bluntly pointed and larger than those following. Very shallow medioventral groove.
Anterior maxillary and dentary teeth conical, unicuspid, with no or very feeble recurvature, becoming larger posteriorly.
The left hemipenis ( Fig. 5 View Fig ) extends about four subcaudal rows when adpressed to the tail, and is about 7.0 mm in length and 4.0 mm across its widest point when fully everted (organ not maximally expanded because one of the lobes was slightly damaged during the eversion process). Hemipenial body globose, tapered near the base, weakly and symmetrically bilobed. Sulcus spermaticus broader near crotch, relatively shallow, curving about halfway around the base of the organ, then running medially towards the lobes. Sulcus flanked by a narrow nude area on each side, and divided in the crotch by a small fleshy bumplike structure, from where two narrow, ill-defined and barely detectable sulcus branches seem to extend laterally into the folded thickened 'ear-like' tissues of each lobe towards the apices of the lobes. Lobes are complexly folded, not flattened terminally, and sulcus branches seem to end in small depressions (from which residual seminal fluid was removed before hemipenis preparation).
Very base of hemipenis nude, pigmented in life. Lateral and asulcate faces of the organ with a series of roughly equidistant oblique plicae or flounces with weakly calcified spinules. Twenty-three rows of flounces extending along the organ. Basal flounces shorter, almost in straight transverse lines, the three first rows almost continuous across the hemipenial base, except for narrow nude areas on each lateral face and for nude areas edging each side of the sulcus spermaticus; five subsequent rows continuous across hemipenial body (except for nude areas edging each side of the sulcus spermaticus), with the seventh and eight rows distinctly pointing medially towards the apex on the asulcate face (those eight flounces proximal to the median nude space of the asulcate face correspond to the 'medioproximal asulcate flounces' sensu Myers et al. 2009); fifteen remaining rows chevron-shaped, also circling the organ, but separated by a broad median nude space on the asulcate face and by the nude areas edging each side of the sulcus spermaticus. The plicae bear comblike rows of minute weakly mineralized spinules that project slightly from the edges of the plicae and usually increase in size towards the lateral faces and the basal part of the organ (staining of the spinules by Alizarin Red is well visible under the microscope although not observable in Fig. 5 View Fig ).
Colour of the Holotype in life
Dorsum greyish to bluish brown depending on light intensity, devoid of any conspicuous pattern, but with a few scattered black scales on flanks. Arms and legs, including digits, greyish brown with a few scattered black scales; rear of thighs with a few ill-defined dark markings. Ventral surfaces of head and body golden grey, ventral surfaces of legs and tail bluish grey, palms and soles lemon yellow speckled with brown and yellowish orange scales. Tympanum dark brown, iris pale yellow ( Figs 1A, C View Fig , 4).
Colour of the Holotype in preservative
After approximately two months in preservative, the dorsal surfaces became dark brown (looking slightly peppered when specimen is in liquid), the ventral surfaces are metallic blue, except the anterior surface of head, which is light grey ( Fig. 6 View Fig ). Palms and soles are grey. Tympanum is dark brown. It is worthwhile to note that the ventral skin, and some parts that are light grey (like the anterior underside of head), turn blue or bluer when pressure is applied to the skin.
Variation in paratypes
The paratypes consist in 1 adult ♂ (SVL 60.4 mm), and 2 juvenile ♂ (SVL from 38.2 to 42.6 mm). Table 1 provides measurements of the type series, and Fig. 1B View Fig shows one of the juvenile paratypes in life (IRSNB 2675).
One of the main differences among specimens of the type series is the occurrence of a presupraocular scale in the three paratypes (absent in the holotype); that presupraocular scale is divided on both sides in IRSNB 2675. IRSNB 2676 has four suboculars on both sides (vs. three in the holotype and other paratypes). Supralabial counts are consistent, but infralabial counts vary from 5 (n=1) to 7 (n=1, one side only). Pits on head scales are inconspicuous in the two juveniles (vs. well visible in the adult males). Suboculars are always separated from palpebrals by 2-3 poorly defined rows of inconspicuous tiny scales. In IRSNB 2674 three of these scales (vs. two in the holotype) are distinctly enlarged (larger than in holotype) with angular downward protrusion between suboculars. In both juvenile paratypes, only one of these scales is only slightly enlarged and slightly protrudes between suboculars. The condition of the parietals and interparietal is highly variable among specimens and none of the specimens examined has complete interparietal and parietals, which are always divided in several smaller scales (see Fig. 7 View Fig for comparison). Number of middorsal scales varies from 53 to 57. Ventrals are in 11 to 14 longitudinal rows and in 29 to 30 transverse rows. The number of femoral pores varies from 8 to 10. Preanals vary from 7 to 10.
The fully everted, maximally expanded, right hemipenis of the adult ♂ paratype (IRSNB 2674) is identical to the holotype's organ, except that maximal expansion makes the sulcus spermaticus bifurcation and the sulcus branches even more difficult to appreciate. Maximal expansion does not highlight hemipenial features that would have been overlooked in the not maximally expanded hemipenis of the holotype. The hemipenis is 6.6 mm in length and 4.0 mm across its widest point when everted; there are twentyfour rows of flounces extending along the organ, ca. eight of which are medioproximal asulcate flounces.
Colour in life ranges from beige to dark greyish, sometimes bluish, brown dorsally, depending on light intensity. Rear of thighs usually has a few ill-defined dark markings. Some blue markings may be visible around ear, or at the base of the tail, or on ventral parts. Ventral parts range from golden grey to pale yellow. One of the juvenile paratypes (IRSNB 2675, see Fig. 1B View Fig ) has two poorly defined dark brown stripes between eye and ear. In preservative all specimens became dark brown dorsally, looking slightly peppered when in liquid. Ventral parts are always metallic blue ( Fig. 6 View Fig ). Tympanum in preserved specimens varies from dark brown to blue or golden blue.
Distribution and ecology
The new species is known only from the Chimantá Massif in Venezuela ( Fig. 8 View Fig ) where it has been reported from Abakapá-tepui (type locality), Amurí-tepui, and Murei-tepui (sometimes named Eruodatepui, see Discussion) between ca. 2100-2600 m elevation ( McDiarmid & Donnelly 2005). The species is probably widespread in the Chimantá Massif.
Anadia mcdiarmidi is diurnal and inhabits open, mostly flat sandstone areas on tepui summits ( Fig. 1D View Fig ). The holotype and two paratypes were collected during the day under rocks; IRSNB 2674 was found during the day, crawling in a small crack between rocks. Two white fertile eggs (both measuring 12.3 mm) and several older eggshells attributable to the new species were found under a large rock, suggesting that a same suitable nesting site may be reused [as already postulated by Gorzula (1992)], and that females lay two eggs at a time. The only other lizard found syntopic with Anadia mcdiarmidi was Neusticurus cf. rudis , which was also mainly found under rocks.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anadia mcdiarmidi
| Kok, Philippe J. R. & Rivas, Gilson A. 2011 |
Anadia breweri
| Brewer-Carias C. & Audy M. 2010: 210 |
Anadia
| McDiarmid R. W. & Donnelly M. A. 2005: 514 |
Anadia
| Gorzula S. & Senaris J. C. 1999: 114 |
Anadia breweri
| Gorzula S. 1992: 276 |