
Protocupressinoxylon dragastanii Iamandei and Iamandei, 2000
|
publication ID |
https://doi.org/10.35463/j.apr.2022.01.04 |
|
DOI |
https://doi.org/10.5281/zenodo.10975574 |
|
persistent identifier |
https://treatment.plazi.org/id/34188785-1D4D-FFBD-94DB-E455FE61582D |
|
treatment provided by |
Felipe |
|
scientific name |
Protocupressinoxylon dragastanii Iamandei and Iamandei, 2000 |
| status |
|
Protocupressinoxylon dragastanii Iamandei and Iamandei, 2000
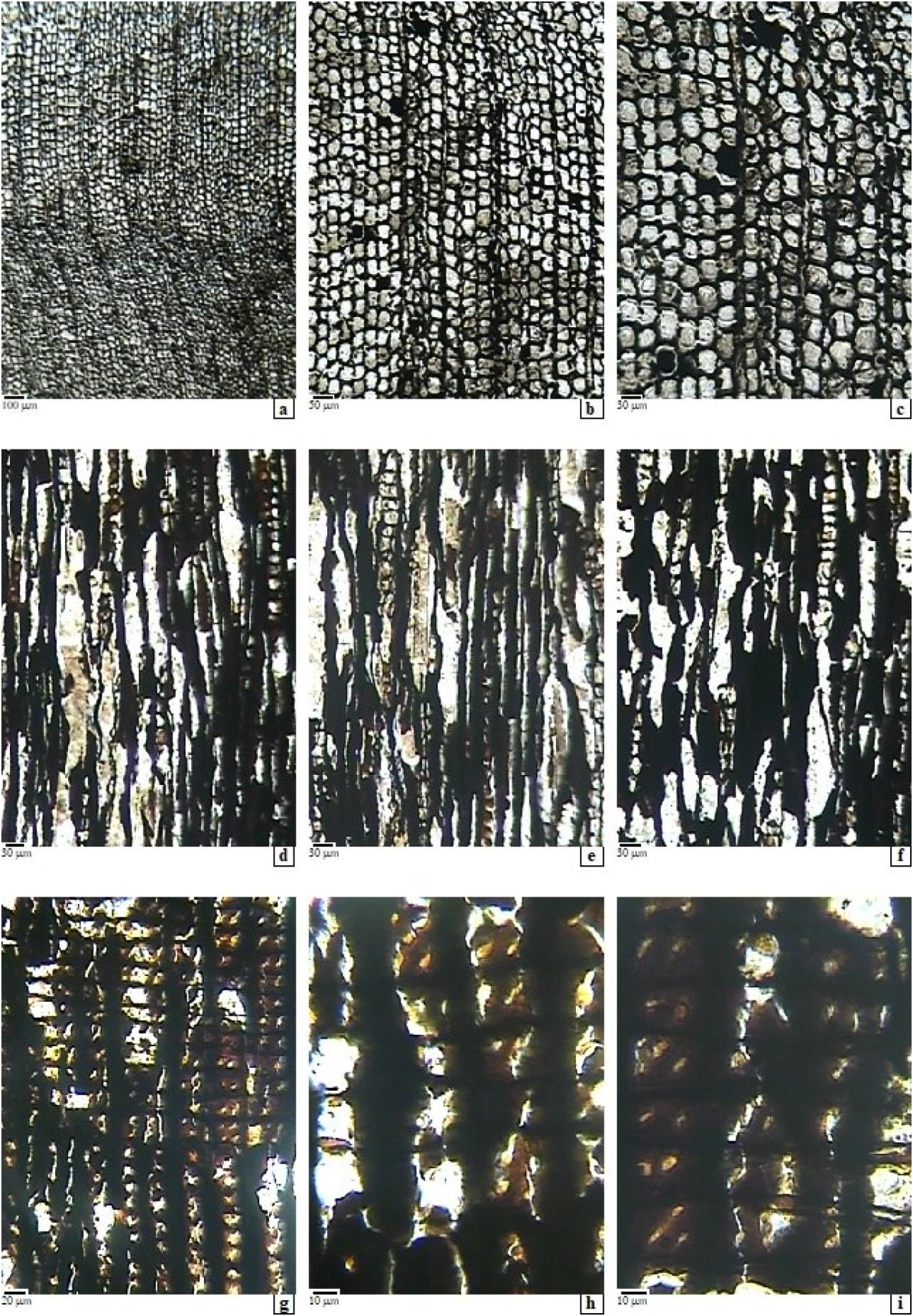
Fig. 1 View Fig , a-i.
Material
The studied material is represented by 14 samples of silicified wood collected from the Holbav area. All studied samples are fragments of trunk or thick-branches, of decimetric size, dark to black color and by magnifying glass or even by naked eye, the regular fibrous structure without vessels suggesting a conifer wood. The described specimens attributed to this taxon have very similar details and have the following field-numbers within the "Grădinaru Collection": 1074, 1077, 1082, 1084, 1086, 1088, 1092, 1097, 1098, 1099, 1100, 1104, 1106, 1109 and are stored at the National Museum of Geology, in Bucharest, under the inventory numbers: 27691, 27692, 27693, 27694, 27695, 27696, 27697, 27698, 27699, 27700, 27701, 27702, 277203, 27720.
Microscopic description
Growth rings – in the studied samples represent secondary xylem, which is tracheidoxylic. Thus, in cross-section they appear more or less well developed, sometimes wide with more than 60 cells, showing a fairly abrupt or gradual transition from early- to late-wood and distinct boundaries of the growth-rings. Normal or traumatic axial resin canals absent.
Tracheids – in cross-section appear large and with relatively thin walls in the early-wood, having quasi-polygonal cross-section (4-5 sides), and are sometimes deformed by compression. They have radial/tangential diameters of 30-60(70) / 25-60(70) μm and the wall thickness of 4-7 μm double wall. Where gradual size transition is present, a well-developed transitional wood is visible, with tracheids gradually diminishing their section and tracheidal wall becoming progressively thicker, until the final 3-5 rows of the late-wood, which consist of smaller radially flattened cells (with r/tg diameters of 20- 25 / 10-20 μm) and thicker walled, of 8-10 μm across the double-wall, marking the growth-ring boundary. Between two successive rays, 1-9 radial rows of tracheids can be counted. Sometimes, radial rows of smaller-sized tracheids are intermingled. The density is between 750-1216 tracheids / mm 2. In longitudinal view, tracheids have on the radial walls 1-2-seriate pits, sometimes contiguous, and of mixed type, because when are biseriate the pits are often alternately arranged. Sometimes crassulae are present. The pits are of abietinean type, have 10-12 μm diameter and round to oval apertures, of 5-6 μm. No details about torus. On the tangential walls, pits are usually absent, or rarely present, as poorly preserved, uniseriate, small round pits, of 5-6 μm in diameter, with small round to oval aperture. Inside the tracheids, organic deposits are sometimes present as dark remains. Average tracheid length is difficult to measure. Helical thickenings on tracheids are absent.
Axial parenchyma – in the cross sections is usually absent or rarely present, as dispersed thin-walled cells with dark content. In longitudinal view it appears as rows of rectangular cells, with the horizontal wall thin and smooth, rarely feebly nodular and with some dark remains inside.
Rays – usually appear thin in cross section while in tangential sections appear exclusively uniseriate, sometimes with some biseriate storeys, having up to 15 cells in height, sometimes more. Their density is 6-9 rays per tangential millimeter. In radial view the rays show homocellular aspect, with parenchymal cells all procumbent, 18-20 μm tall, marginals slightly taller, of 25-30 μm. The horizontal ray-cell walls are smooth or slightly rough and usually pitted, and are relatively thin (2-3.5 μm double wall); the tangential end-walls are relatively thick, of 1.5- 2 μm the simple wall, smooth or slightly nodular, but usually poorly preserved. The indentures are missing or difficult to notice. Spiral thickenings were not observed, most probably are absent. The cross-field pitting is of cupressoid type, showing 1-2(4-5) pits, in 1-2 rows horizontally arranged, and sometimes irregular or even to slightly alternate aspect. The pits are oculipores of cupressoid type, 8-10 μm in diameter, with slit- like or lenslike apertures of 2-5 μm. Sometimes cross- field pitting is difficult to observe, due to poor preservation or to the presence of organic content.
Radial or axial resin canals – are absent.
Mineral inclusions – usually are not present.
Affinities and discussion
The xylotomy of the here studied 14 specimens is definitely pycnoxylic and homoxylic (or tracheidoxylic – sensu Creber, 1972), having tracheids with radial pitting of mixed type, no resin canals, little or no parenchyma, exclusively uniseriate rays and cross-fields with 1-2(4-5) cupressoid pits with slit-like inclined apertures, arranged in one or two horizontal rows, which are slightly irregularly to even alternately arranged; all these characters are evidence of a typical cupressaceous structure. Such details as: radial pitting on tracheids of mixed type, cupressoid pits in cross-fields, to fairly araucarioid arrangement, axial parenchyma showing few and radial pitting of tracheids of mixed type in a Mesozoic wood structure indicate a wood-type from “ Cheirolepidiaceae family”, as defined by Alvin et al. (1981) and Alvin (1982).
The synthetic description of all the studied specimens was compared with the diagnoses of Mesozoic genera with cupressoid oculipores in the cross-fields as described by Phillips (1941), Kräusel (1949), Vogellehner (1967, 1968), Müller Stoll and Schultze-Motel (1989), Philippe (1995, 2002), Bodnar (2017), and it appears that our material is mostly consistent, at generic level, with Protocupressinoxylon wood-type. In fact, the Mesozoic conifers were grouped, a long time ago, based on anatomical features, in the " Protopinaceae family" ( Krausel, 1917 This family name was noted by Bamford et al. (2016) as having a dubious taxonomic status, and they recommended its abandon.
However, the name Cheirolepidiaceae , attributed to Turutanova-Ketova (see Fossil works – online), is not a valid name, as remarked by Doweld (2020), who considered an alternative name as being more correct for the Mesozoic gymnosperms characterized by pollen of Classopollis - type. Thus, he applied the valid family name Hirmeriellaceae Harris, 1979 - and proposed this family name “for conservation against Cheirolepidiaceae ”, the latter being considered as "inadmissible, being based on the illegitimate later homonym Cheirolepis Schimper , and thus, itself illegitimate” (see Doweld, 2020).
This genus name, Protocupressinoxylon , was established by Eckhold (1922, pp. 490-491), to designate Mesozoic fossil conifer wood-types described like this: “wood with annual rings more or less distinct, tracheid pitting in various transitional arrangements, both horizontal and end walls of ray parenchyma cells smooth, resin canals absent, axial parenchyma occasionally present, cross-field pits cupressoid”, but not specifying their number and arrangement.
Initially, Eckhold (1922) included seven species in the protologue: Protocupressinoxylon cupressoides , P. jurassicum , P. potomacense , P. eboracense , P. koettlitzi , P. vectense and P. mesozoicum , but without designation of a clear generotype. Only later, Andrews (1955) designated Protocupressinoxylon cupressoides (Holden) Eckhold, 1922 (not 1923, see Jogmans and Dijkstra, 1974, p. 794) - as the species-type for that generic name. However, the species Protocupressinoxylon eboracense which was initially assigned as type-species of the Protobrachyoxylon genus ( Holden, 1913) was included by Eckhold (1921) along with other species in the protologue of Protocupressinoxylon genus. But the original material seems to be lost, and the analysis of Holden’s description and figuration suggest either identity with Protocupressinoxylon Eckhold, 1921 or with Brachyoxylon Hollick and Jeffrey, 1909 (see Bamford and Philippe 2001, Philippe, 2002).
Recently, Bodnar (2017) proposed the genus name Protocupressinoxylon to be conserved, together with a proposed generotype – P. malayense Roggeveen, 1932 . Also, she proposed the necessity of a systematic revision of several described taxa, since abnormal details were described for some species of Kräusel (1949), like resin ducts in Protocupressinoxylon potomacense , P. solmsi , P. vectense , P. luccombense , or multiseriate rays, like in P. weidlingense (in Kräusel, 1949, pp. 183-185).
For this genus many species of fossil wood from all over the world ranging from Permian to Cretaceous have been described, especially after Kräusel (1949), who admited it as valid genus, and he even has described two species: P. dockumense (Torrey) Kräusel and P. coromandelinum (Sahni) Kräusel.
Later, new European species were described, as for example: Protocupressinoxylon catenatum Schultze-Motel 1960 (see also Iamandei and Iamandei, 2001), P. liasinum Schultze-Motel 1960 , P. leonardianum (Charrier) Vogellehner 1968 , P. rhaeticum Vogellehner 1968 , P. munense Shilkina and Blokhina (in Blokhina, 1975), P. purbeckensis Francis 1983 , P. carentanensis Barale (in Philippe, 1995).
Also, Iamandei and Iamandei (2000, 2005) have described an Early Cretaceous species from Dobrogea, Romania: Protocupressinoxylon dragastanii Iamandei et Iamandei , which has radial pitting uni- to biseriate, opposite or alternate, no parenchyma, rays of mid tall, uniseriate or partially biseriate and cross fields of cupressoid type, sometimes with an araucarioid tendency (i.e. alternate pits).
More recently, Correa et al. (2018) studying an abundant population of wood remains (58 permineralized trunks) from Argentina, described a new species as Protocupressinoxylon carrizalense , and made a comparative table of 32 species. They observed that only 24 from them fit the generic diagnosis, but eight species (i.e. Protocupressinoxylon alternans , P. cenomanicum , P. mesozoicum , P. luccombense , P. jurassicum , P. koetlizi , P. potomacense and P. vectense ) present cross-fields pits that are different from the cupressoid type: either not bordered, or typical taxodioid (see Correa et al., 2018).
Since the specific details inside the genus Protocupressinoxylon are very similar, it is very difficult to discriminate between them, but through comparative study of their descriptions, and using also the comparative table from Correa et al. (2018), we tried to identify our material, which present these clear xylotomical features: tracheidoxylic secondary wood, having radial pitting 1-2 seriate, of mixed type on tracheids, few parenchyma or absent and uniseriate rays with cross-fields with 1-2(4-5) cupressoid pits per field, spaced arranged in 1-2 horizontal rows when more, to irregular, or to slightly alternate. These xylotomical details are very similar with those described by Iamandei and Iamandei (2000 and 2005) for the species Protocupressinoxylon dragastanii from the Early Cretaceous of South Dobrogea, Romania.
Thus, based on the above discussion, we attribute all the 14 studied specimens to the species Protocupressinoxylon dragastanii Iamandei and Iamandei, 2000 , as another Mesozoic European taxon.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |