
Rhamdella cainguae, Bockmann, Flávio A. & Miquelarena, Amalia M., 2008
|
publication ID |
https://doi.org/10.5281/zenodo.182293 |
|
DOI |
https://doi.org/10.5281/zenodo.5672224 |
|
persistent identifier |
https://treatment.plazi.org/id/36371172-B462-0967-FF7C-F9204668B53E |
|
treatment provided by |
Plazi |
|
scientific name |
Rhamdella cainguae |
| status |
sp. nov. |
Rhamdella cainguae View in CoL new species
( Figs. 1 View FIGURE 1 and 2 View FIGURE 2 )
Rhamdella sp.— López et al. 2003 (citation: 63).
Holotype. ILPLA 1078, 130.0 mm SL, male, Argentina, Provincia Misiones, Departamento Cainguás, Aristóbulo del Valle, Arroyo Cuña-Pirú in the Balneario Municipal ( 27º10´S 54º50´W), coll. F. de Durana, R. Filiberto and H. Oñatibia, 22–26.ix.1997.
Paratypes. ILPLA 468, 2 males in alcohol (115.0–140.0 mm SL) and 1 male C & S (114.0 mm SL), same locality and collectors as holotype, 20.vii.1996; ILPLA 1019, 4 males (116.0–125.0 mm SL), 5 females (100.0– 126.8 mm SL mm SL), 1 indet. (88.0 mm SL), 1 male C & S (138.0 mm SL), LIRP 3045, 1 male in alcohol ( 153.8 mm SL), 1 female in alcohol ( 107.2 mm SL), and 1 male C & S ( 149.9 mm SL), MLP 9530, 1 male (129.0 mm SL), MNRJ 23157, 1 male ( 153.2 mm SL), MZUSP 81427, 1 female ( 124.6 mm SL), same locality and collectors as holotype, 18–22.ix.1997; ILPLA 1079, 3 males ( 113.9–157.5 mm SL), same locality as holotype, coll. L. Alcalde and R. Filiberto, 25–27.vii.1998; ILPLA 1354, 1 male ( 122.4mm SL) and 2 females ( 100.7–107.7 mm SL), same locality as holotype, coll. R. Filiberto, H. López and E. Etcheverry, 18.iii.2000; ILPLA 1176, 2 males ( 148.7–153.4 mm SL), Argentina, Provincia Misiones, Departamento Cainguás, Aristóbulo del Valle, Arroyo Tateto, on road to La Misión, coll. R. Filiberto, A. Miquelarena and M. Montenegro, 18–22.ix.2000.
Diagnosis. Rhamdella cainguae is autapomorphically diagnosed by having a distinct and large ovoid area in the supraorbital laterosensory canal between the frontal and sphenotic delimited by the apparently slender dorsal walls of these bones and with no foramen for a laterosensory branch. The new species is further distinguished from congeners by the following combination of characters: from R. aymarae Miquelarena and Menni 1999 , by a shorter maxillary barbel (barbel tip reaching from the base of pectoral-fin ray in a 88.0 mm SL specimen, to the posterior portion of opercular region, not surpassing the branchial slit, in a 157.5 mm SL specimen vs. tip reaching to vertical through close to the dorsal-fin base in a 90.0 mm SL specimen, to the vertical through the dorsal-fin origin in a 150.0 mm SL specimen), larger horizontal eye diameter (20.6–23.9% of HL vs. 11.5–17.3%), lesser interorbital distance (17.9–20.7% of HL vs. 30.0–44.9%), snout profile slightly convex (vs. snout profile distinctly straight), more numerous branchiostegal rays (seven vs. six), branchiostegal membranes not reaching the basal part of the first pectoral-fin ray in most specimens (vs. extending over the basal part of first pectoral-fin ray in most specimens), shorter interdorsal length (7.6–11.3% of SL vs. 30.0–44.9%), longer adipose-fin base (36.0–41.6% of SL vs. 18.5–27.9%), longer anal-fin base (16.6–22.0% of SL vs. 12.3–17.8%), more numerous anal-fin rays (16–18, commonly 17, vs. 15–16); and dorsal caudal-fin lobe much longer than ventral lobe (length of ventral caudal-fin lobe is 55.8–65.5% of length of dorsal lobe in males vs. 81.4–94.3%); from R. eriarcha ( Eigenmann and Eigenmann 1888) by the dorsal caudal-fin lobe much longer than ventral lobe (length of ventral caudal-fin lobe 55.8–65.5% of length of dorsal lobe in males vs. 75.0–94.7%); from R. longiuscula Lucena and Silva 1991 , by the smaller horizontal eye diameter (20.6– 23.9% of HL vs. 22.5–29.4%), larger interorbital distance (17.9–20.7% of HL vs. 12.8–17.7%), greater snout length (37.9–43.2% of HL vs. 28.3–37.5%), snout profile slightly convex but almost straight (vs. snout profile distinctly convex), and branchiostegal membranes not reaching the basal part of the first pectoral-fin ray in most specimens (vs. extending over the basal part of the first pectoral-fin ray); and from R. rusbyi Pearson 1924 , by shorter maxillary barbels (their tips reaching from the base of the pectoral-fin ray in a 88.0 mm SL specimen, to the posterior portion of opercular region, but not surpassing the branchial slit in a 157.5 mm SL specimen vs. tip reaching to the vertical through the posterior end of the dorsal-fin base or slightly surpassing that point in specimens of 134.2 to 183.8 mm SL), smaller interorbital distance (17.9–20.7% of HL vs. 26.5– 27.1%), higher number of branchiostegal rays (seven vs. six), shorter pelvic fin (13.3–16.9% of SL vs. 17.0– 19.1%), longer anal-fin base (16.6–22.0% of SL vs. 11.8–14.0%), higher number of anal-fin rays (16–18 analfin rays, commonly 17, vs. 12–14); and dorsal caudal-fin lobe much longer than ventral lobe (length of ventral caudal-fin lobe 55.8–65.5% of length of dorsal lobe in males vs. 84.8–93.7%); a dense concentration of long and slender papillae on the lateral body surface in the pectoral and abdominal regions, most conspicuous near the lateral line, resulting in a hairy aspect (vs. papillae comparatively much longer and larger, especially proximate to lateral line); and by the presence of a distinct narrow dark mid lateral stripe (vs. stripe absent).
Remarks. Miquelarena and Menni (1999) utilized the degree of ossification and branching of the lateral line as distinguishing features between R. rusbyi and R. aymarae . The degree of ossification and branching of the lateral line of R. rusbyi , with well-developed ossicles extending posterior to the middle of caudal fin, is the most extreme condition among Rhamdella species, where ossification of the lateral line otherwise reaches only to the level of the anal fin. However, those characters were observed in only one specimen of R. rusbyi (CAS 63729), which is larger than any C & S specimen of other Rhamdella species. Since large individuals tend to have a higher degree of ossification and branching of the laterosensory system than small ones, the degree of ossification and branching of the lateral line of R. rusbyi may be due to large size only.
Holotype Paratypes
Sex male 10 males 5 females
Range Mean SD Range Mean SD
Standard length (mm) 130 113.9–157.5 100–126.8
Percentage of standard length
Body depth 17.7 14.6–19.0 16.8 1.5 16.6–19.5 18.0 1.2 Body width 16.9 17.0–19.5 17.9 0.7 18.4–20.7 19.2 1.0 Head length 26.1 25.8–28.8 26.7 0.9 27.4–29.2 28.4 0.8 Head width 17.7 17.0–19.5 18.3 0.8 18.9–21.0 20.0 0.8 Prepectoral length 24.3 24.0–28.0 25.6 1.2 26.0–29.4 27.9 1.3 Predorsal length 34.4 33.3–38.4 36.0 1.6 37.3–40.4 39.2 1.3 Preventral length 47.7 45.7–50.4 48.5 1.3 48.7–53.3 51.2 2.2 Preanal length 60.8 60.0–64.3 62.4 1.5 63.9–68.4 66.2 1.6 Preadipose length 52.8 53.3–57.7 55.9 1.7 57.6–60.6 58.9 1.2 Length of pectoral spine 13.1 11.9–14.0 12.9 0.7 13.1–14.1 0.4 Length of dorsal spine 11.1 9.5–13.2 11.4 1.4 9.5–11.8 10.5 0.9 Length of ventral fin 14.6 13.3–16.9 14.9 1.2 15.0–16.0 15.5 0.5 Dorsal-fin base 14.6 13.3–16.5 14.5 1.1 12.5–14.6 13.9 1.0 Anal-fin base 19.5 18.6–22.0 20.3 1.1 16.6–19.3 18.1 1.2 Adipose-fin base 39.7 37.5–41.6 39.6 1.3 36.0–41.5 38.2 2.1 Caudal peduncle length 22.0 21.2–24.6 22.5 1.0 21.7–23.6 22.8 0.7 Caudal peduncle height 9.0 8.5–10.1 9.1 0.6 9.0–9.8 9.3 0.4 Adipose height 4.6 4.1–5.3 4.5 0.4 4.2–5.6 4.8 0.5 Distance dorsal-adipose 8.8 7.6–10.3 8.9 0.9 8.5–11.3 9.8 1.2 Urogen. papilla-anal-fin origin 2.8 2.6–3.5 3.1 0.3 4.7–5.1 4.9 0.2 Description. Morphometric data are given in Table 1. Cross-section of trunk roughly elliptical at level of dorsal-fin origin, gradually more compressed posterior to dorsal-fin base. Caudal peduncle region compressed, merging with caudal fin from dorsal view. Anterodorsal profile of body gently convex, curving in an even arch from snout to base of dorsal fin, except for discrete depression at base of supra-occipital process. Dorsal profile gently concave to straight from dorsal-fin base to adipose-fin origin and approximately straight or gently convex from that point to base of caudal fin. Ventral body profile in a continuous convex curve from mouth to pelvic-fin base. Ventral profile of body straight from pelvic-fin insertion to caudal-fin base. Fortyone to 43 myotomes apparent through skin; myotomes becoming progressively narrower and more angled posteriorly. No deposits of fatty material in globose bodies, commonly seen in heptapterids, perceptible through the skin. Pectoral pore absent. Anus and urogenital pore very close to each other.
Skin of top and underside of head, as well as opercular and branchiostegal regions with dense covering of papillae; lateral surface of the pectoral and abdominal regions, approximately to vertical through posterior end of dorsal-fin base with long, slender papillae that give a hairy aspect to that region; papillae most concentrated near lateral line. Skin of ventral surface of body apparently without papillae.
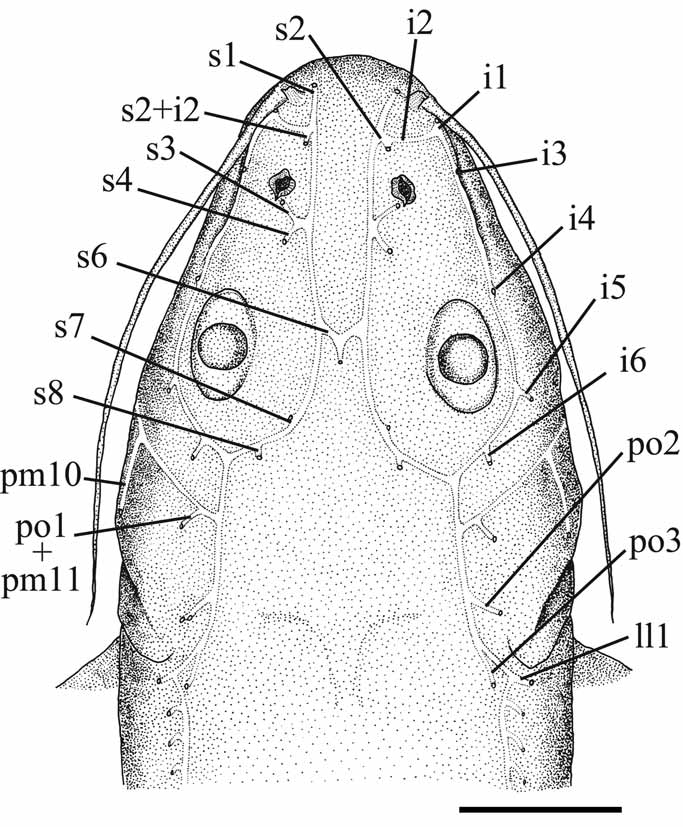
Head long (greater than 25% of SL), narrow (its greatest width about 0.7–0.8 times its length), deep (its greatest depth nearly 0.5–0.6 times its length), and elliptical in dorsal view ( Figs. 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). Deep longitudinal facial ridge marking dorsal limits of adductor mandibulae muscle, extending from base of maxillary barbel to or just anterior of level of fourth infraorbital pore ( i4). Distance between posterior nares slightly greater than that between anterior nares. Anterior and posterior nares far apart, with separation slightly less than distance between anterior and posterior nares. Four nares arranged as in vertices of an square. Anterior nare tubular with lateral or posterolateral limit adjacent to anteriormost supraorbital pore (s1) and anteromedial to anteriormost infraorbital pore ( i1). Anterior nare close to upper lip. Posterior nare rounded, situated immediately anterior to third supraorbital pore (s3), with high, anteriorly deepest, raised margin, surrounding it almost completely, other than at posterolateral point. Posterior nares about midway between anterior nares and posterior margin of eye. Posterior nare lacking barbel. Adductor mandibulae muscles restricted to facial region, accommodated on lateral wall of hyomandibula, and not extending dorsally to midline of head. Levator operculi muscle inserted on the dorsal and dorsoposterior portion of opercle.
Mouth wide, subterminal, with snout projecting slightly beyond jaw ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ). Gape transverse, slightly ventrally curved at corners, with fleshy rictal fold at corners. Rictal fold well developed, ventrally subtended by submandibular groove that extends dorsally around corner of mouth posterior to upper part of rictal fold and ventrally to point between second and third preoperculomandibular pores (pm2 and pm3). Both upper and lower lips subdivided by a deep depression into two transverse folds; with several longitudinal plicae. Premaxillae, dentaries and pharyngeal tooth plates with small villiform teeth arranged in bands. Toothbearing portion of premaxillae long and narrow, rectangular, with projections onto posterolateral angle of bone, and usually partially in contact at symphysis. Dentary tooth patch slender and posteriorly elongate. Palate edentulous. Upper and lower oral valves thin, covered by papillae, with posterior margins concave. Upper oral valve totally free posteriorly. Lower oral valve broadly attached anteriorly to mouth floor just behind dentary dentition, and with free rounded flaps posteriorly. Upper valve approximately as long as upper tooth plate, its length decreasing progressively posteriorly; lower valve narrower anteriorly and progressively wider posteriorly, forming large posterior flaps. Mouth roof and floor with sparse papillae.
Barbels short, slender, and tapering progressively towards their distal extremities ( Figs. 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). Barbels approximately ovoid in cross section, without fringing membrane. Maxillary barbel, longest, inserts above upper lip lateral to anterior nare and adjacent to anteriormost pore of infraorbital laterosensory canal ( i1). Tip of adpressed maxillary barbel reaches branchiostegal membrane (but not surpassing the branchial slit) in larger specimens; and reaches posteriormost third of first pectoral-fin ray, in advance of dorsal-fin origin, in smaller specimens. Mental-barbel base inserted midway between anterior border of lower jaw and gular fold. Outer mental barbel longer than inner barbel, and inserted nearly behind fourth pore of preoperculomandibular laterosensory canal (pm4). Tip of adpressed outer mental barbel reaches from point slightly posterior to the branchiostegal fold to point slightly beyond outer border of branchiostegal membrane. Inner mental barbel inserted slightly in advance of vertical through origin of outer mental barbel, approximately behind second pore of preoperculomandibular laterosensory canal (pm2). Tip of adpressed inner mental barbel reaches from point just posterior to branchiostegal fold to point almost at outer border of branchiostegal membrane.
Gular fold distinct, fleshy, and broadly V-shaped with rounded apex ( Fig. 5 View FIGURE 5 ). Posteroventral portion of opercle and branchiostegal region delimited by a distinct ridge on lateral surface of head extending from distal tip of opercle to a point between eighth and ninth pores of preoperculomandibular canal (pm8 and pm9), ventrally.
Branchiostegal membranes well developed, thick, united to isthmus only at medial apex, and firmly connected to each other anteriorly ( Fig. 5 View FIGURE 5 ). Branchiostegal membrane not reaching base of first pectoral-fin ray and not overlapping ventrally. Gill rakers thick and moderately long, with: 4 (1), 5 (1), 6 (6*), 7 (3), or 9 (1) rakers on first ceratobranchial, and 1 (3), 2 (7), or 3 (2*) on first epibranchial, plus 1 (12*) on angle formed by these bones.
Eyes large , elliptical (greatest length in longitudinal axis), with rims circumscribed with deep, continuous invagination, distinctly more pronounced at anterior and dorsal borders ( Figs. 3 View FIGURE 3 and 4 View FIGURE 4 ). Skin over eye thin and transparent, with lens clearly visible. Eye dorsolaterally positioned, centered approximately at midpoint between tip of snout and corner of opercular membrane and situated proximate to each other, being separated by a space slightly larger than longitudinal diameter of eye. Pupil rounded.
Dorsal fin distally rounded in profile, reaching to adipose fin when adpressed. First dorsal-fin ray (spinelet) very small (not externally visible), triangular and with two ventral limbs, followed by 1 long unbranched, and 6 (10*) or 7 (2) branched rays. Second dorsal-fin ray with most of proximal portion stiffened (54.2–67.4% of entire length) and with segmentation perceptible solely distally, forming delicate spine, and shorter distal portion flexible and clearly segmented. Spiny portion of the second dorsal-fin ray straight, approximately ovoid in cross-section, and lacking marginal dentations. Second dorsal-fin ray slightly shorter than third and fourth rays (first and second branched rays, respectively), whose tips project beyond tip of second dorsal-fin ray. Origin of dorsal fin approximately on vertical through the posterior portion of mid third or anterior portion of most posterior third of adpressed pectoral fin.
Pectoral fin with distal margin slightly convex, with 1 unbranched and 8 (3), 9 (6*), or 10 (3) branched rays. First pectoral-fin ray covered by thick integument; convex anteriorly and straight to slightly concave dorsally. Most of proximal part of first ray rigid and with hardly perceptible segmentation (72.0–82.6% of its entire length), forming a distinct spine; shorter distal portion flexible and clearly segmented. First pectoral-fin ray approximately as long as second (first branched) and third (second branched) rays, but tips of second and third rays projecting slightly beyond the tip of first ray. Spiny part of first pectoral-fin ray with 11–17 conspicuous, regularly spaced dentations along posterior margin, and 26–46 dentations smaller than posterior dentations along its anterior margin (88.0– 153.4 mm SL). There is no evidence of sexual or ontogenetic co-relation with the number of dentations. Pectoral fin lying parallel to main body axis when expanded and slightly directed upwards when adpressed to body.
Pelvic fin convex posteriorly, with 6 rays (12*). Lateral ray unbranched, completely flexible, segmented, and distinctly shorter than second and third rays (first and second branched rays, respectively). Tip of pelvic fin reaches vertical through anal-fin origin. Pelvic-fin origin approximately at or slightly in advance of midpoint of body (excluding caudal fin), and on vertical through base of last dorsal-fin ray.
Anal-fin margin rounded in lateral profile, with 16 (2), 17 (5*), or 18 (1) rays, including 10 (3) or 11 (5*) branched rays: vi+10 (1), v+11 (1), vii+10 (1), vi+11 (4*), or viii+10 (1). Anal-fin base medium-sized; covered basally with thick muscular tissue. Origin of anal-fin base approximately on vertical through anterior fifth of adipose-fin base. Posterior limit of anal-fin base approximately at vertical through middle of last twothirds of adipose-fin base.
Adipose fin long, forming ascending elevated curve in lateral profile, with highest point approximately at last third. Adipose fin merging gradually with back anteriorly, its origin difficult to pinpoint. Distance from dorsal fin to adipose fin less than length of dorsal-fin base. Origin of adipose fin anterior to middle of trunk, and approximately on vertical through middle of adpressed pelvic fin and through, or just behind, anus. Posterior adipose-fin base well defined, forming posterior free lobe markedly distinct from anteriormost portion of dorsal caudal-fin fold. Vertical through end of adipose-fin base distinctly posterior of tip of last anal-fin rays.
Caudal fin deeply forked, lobes long and broad, with rounded contours and 7 and 8 branched rays in dorsal and ventral lobes, respectively. Total caudal fin-rays 50 (1), 52 (1), or 55 (1), with 24 (1), 25 (1), 26 (1) rays in dorsal lobe and 26 (1), 27 (1), 29 (1) rays in ventral lobe. Dorsal caudal-fin lobe much longer than ventral one, markedly so in males: length of ventral caudal-fin lobe 55.8–65.5% of length of dorsal lobe in males (9 ex., 121.0– 157.5 mm SL) and 77.8–80.8 % in females (2 ex., 107.2–124.6 mm SL). Membrane uniting dorsal and ventral caudal-fin lobes extending approximately to the extremities of the median rays of the caudal fin.
Color in life. Overall pigmentation varying from uniformly light to dark gray, or spotted (either with light background color and dark irregular blotches or dark background color and lighter irregular blotches), becoming white ventrally. In light-colored specimens fins almost translucent or yellowish with no conspicuous blotches, fin margin dark in some individuals. Dark-colored specimens with all fin rays intensely gray and fin membranes intensely yellow. Dark , narrow stripe extending from posterodorsal margin of opercle to middle caudal-fin rays. All specimens with iridescent yellow and lilac hues. Mental barbels white; maxillary barbels iridescent gray.
Color in alcohol. Upper portion of head and cheeks densely covered by dark pigmentation, becoming gradually less intense ventral to rim of orbit. Head lightly pigmented ventrolaterally and devoid of pigmentation or with scattered melanophores ventrally (at approximately ventral to line of preoperculomandibular canal). Dark ventral pigmentation mostly concentrated around bases of outer mental barbels. Lower lip with dense concentration of dark chromatophores not extending beyond line of mandibular portion of preoperculomandibular laterosensory canal. Regions around head sensory pores devoid of pigmentation. Maxillary barbel darkly pigmented dorsally and unpigmented ventrally. Mental barbel completely unpigmented or with few melanophores at base. No mask across the eyes, as seen in some Brachyrhamdia species (see Lundberg and McDade 1986; Sands 1985). Overall trunk coloration varying from uniform dark brown to light brown, sometimes with irregularly distributed darker blotches ( Figs. 1 View FIGURE 1 and 2 View FIGURE 2 ). Mid-dorsal region of trunk from dorsal-fin origin to adipose-fin origin typically darker. Lateral surface of trunk with a well-defined narrow stripe of constant width along lateral line with about one third of orbital length. Stripe extends from region immediately behind branchiostegal membrane to bases of median caudal fin-rays. Pectoral and abdominal areas unpigmented except for scattered melanophores behind and below base of pectoral-fin rays, and on areas near base of pelvic-fin rays, and medial portion of basipterygia. Ventral midline posterior to pelvic-fin insertion and along anal-fin base, weakly pigmented. No zone behind the head devoid of pigmentation (unpigmented collar), as exhibited by some Brachyglanis , Cetopsorhamdia , Chasmocranus , and Myoglanis species (see Eigenmann 1912; Schultz 1944). Upper portion of trunk lacking unpigmented areas or dark transverse bars, as in several heptapterid genera (e.g. Heptapterus , Imparfinis , Mastiglanis , Rhamdioglanis , Taunayia etc.). No dark vertical bar along base of the caudal-fin rays, as seen in some Brachyrhamdia species (see Sands 1985). Dorsal-fin rays and adjacent areas of interradial membrane densely pigmented with brown. Dorsal surface of pectoral fin with high concentration of brown or gray chromatophores along rays; interradial membrane translucent, dorsally unpigmented, except for areas adjacent to pectoral-fin rays. Ventral surface of pectoral fin unpigmented or with few scattered small melanophores along rays. Dorsal surface of pelvic-fin rays and adjacent interradial membranes densely covered with brown or gray melanophores. Ventral surface of pelvic fin unpigmented or with few scattered, small chromatophores along rays. Adipose fin with intense concentration of brown or gray chromatophores, and with darker line on its outer margin. Anal-fin rays with dark chromatophores but with most of interradial membrane lacking dark pigmentation except for areas adjacent to fin rays. Muscular basal portion of anal fin and region immediately anterior well pigmented. Caudal-fin rays densely brown-pigmented. Interradial caudal-fin membrane devoid of chromatophores.
Sexual dimorphism. Adult females with urogenital papilla close to anus, urogenital papilla and anus equally wide basally, and papilla with small triangular posterior tip. Adult males with urogenital papilla more distant from anus, base of papilla narrower than anus, and papilla with a more developed triangular posterior tip. Distance from urogenital papilla to anal-fin origin greater in females (4.5–5.1 %) vs. males (2.6–3.5 % SL). Dorsal caudal-fin lobe proportionally more developed in males than females.
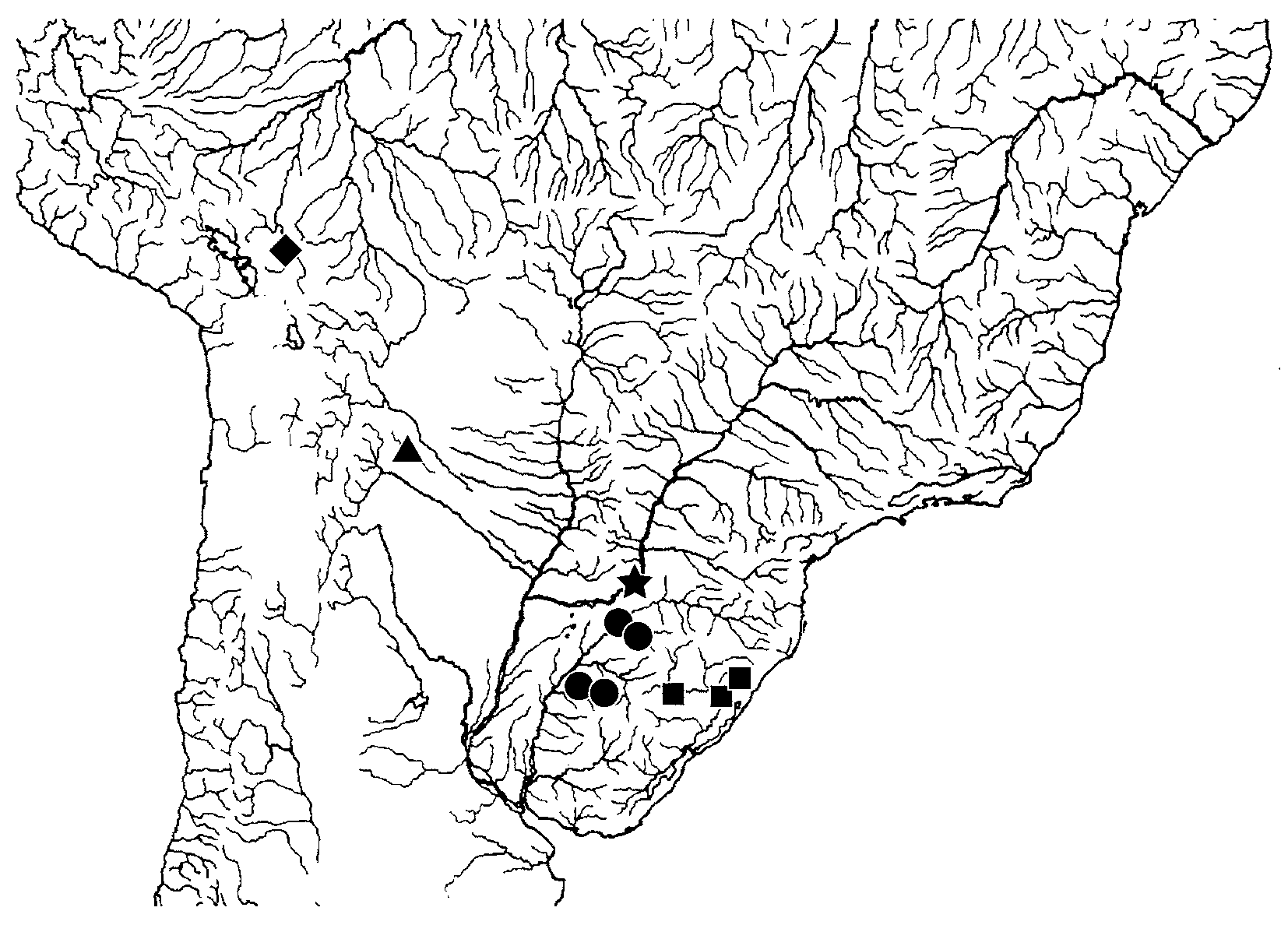
Distribution. Known only from the type locality, Arroyo Cuña-Pirú, Río Paraná basin, in northeastern Argentina ( Fig. 6 View FIGURE 6 ).
Habitat. Specimens were collected with seine or hook-and-line, over rocky bottoms in a clear water stream (ca. 50–100 cm deep, with pools around 3 m deep), running through a densely forested subtropical area. Individuals demostrated a tendency for gregarity as they were densely grouped under large stones.
Etymology. From the Guaraní, ca´á (forest) and iguá (inhabitant), in allusion to the Cainguá, indigenous people of northeastern Argentina.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |