
Cladoxerini Karny, 1923
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4128.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B4D2CD84-8994-4CEF-B647-3539C16B6502 |
|
DOI |
https://doi.org/10.5281/zenodo.6084900 |
|
persistent identifier |
https://treatment.plazi.org/id/387F3068-D32A-FF8A-FF27-ED43260F1C4B |
|
treatment provided by |
Plazi |
|
scientific name |
Cladoxerini Karny, 1923 |
| status |
|
4.2.1. Cladoxerini Karny, 1923 View in CoL
( Figs. 1–5 View FIGURES 1 – 5 )
Type-genus: Cladoxerus St. Fargeau & Audinet-Serville, 1828: 445
Cladoxerinae Karny, 1923: 237 (in part).
Kevan, 1982: 283.
Cladoxerini Günther, 1953: 557 (in part).
Bradley & Galil, 1977: 189.
Otte & Brock, 2005: 32 (in part).
Baculini Günther, 1953: 555 . (Type-genus: Baculum Saussure, 1861 ) n. syn. Zompro, 2004: 134, 135.
Phibalosomatini (Sectio V: Phibalosomata) Redtenbacher, 1908: 399 (in part).
Comments: Karny (1923: 237) originally established the subfamily Cladoxerinae to replace Redtenbacher's Phibalosomini . Günther (1953: 557) first used Cladoxerini for a tribe of his Phibalosominae and Bradley & Galil (1977: 189) listed it as a tribe of Cladomorphinae , while Kevan (1982: 283) treated it as a subfamily of his Bacteriidae . Baculini was originally established by Günther (1953: 555) as a tribe of Phasmatidae : Phasminae (= Phasmatinae Bradley & Galil, 1977) and included exclusively Oriental taxa. Günther's use of Baculini was based on the erroneous interpretation of Baculum Saussure, 1861 by Kirby (1904a: 327), who erroneously synonymised it with the Oriental Clitumnus Stål, 1875 . In fact, Saussure (1861: 127) originally described Baculum as a subgenus of Bacillus Latreille, 1825 , the type-species being the Brazilian Baculum ramosum Saussure, 1861 by monotypy. Kirby (1904a: 327) erroneously selected the Sri Lankan Bacillus cuniculus Westwood, 1859 as the type-species of Baculum , although Saussure had not included this species in the genus. Thus it was not available from the viewpoint of zoological nomenclature and cannot serve as the type-species for Baculum . Hennemann (2002) has partly clarified the confusion around the Oriental taxa erroneously assigned to Baculum and selected the Sri Lankan Bacillus cuniculus Westwood as the type-species of Cuniculina Brunner v. Wattenwyl, 1907, an exclusively Oriental genus. Hence, Hennemann & Conle (2008: 67) re-established Clitumnini Brunner v. Wattenwyl, 1893 for the Oriental genera that Günther (1953: 555) contained in Baculini .
Zompro (2004: 135) discussed the genus Baculum , provided a re-description of the type-species B. ramosum , whose type is lost, and correctly assigned Baculini to the Cladomorphinae . However, Zompro did not identify the close relation between the newly recognized Baculini and Cladoxerini . The species which Zompro (2004: 136, fig. 2) described as Baculum ramosum is without any doubt congeneric with the type-species of Cladoxerus St. Fargeau & Audinet-Serville, 1828 , i.e. C. gracilis St. Fargeau & Audinet-Serville, 1828 . Hence, Baculum Saussure, 1861 falls as a junior synonym of Cladoxerus (n. syn.) and the tribe Baculini consequently becomes a junior synonym of Cladoxerini Karny, 1923 (n. syn.).
The genus Parabactridium Redtenbacher, 1908 (Type-species: Parabactridium mirum Redtenbacher, 1908: 403 ) was described on the basis of ♀♀ from Madagascar in MNHN and NHMW and originally placed in close relation to Cladoxerus St. Fargeau & Audinet-Serville, 1828 . Hence, subsequent authors (e.g. Bradley & Galil, 1977 or Otte & Brock, 2005) have placed the genus in the tribe Cladoxerini . However, the type-specimens appear to be lost and the systematic position within Cladoxerini has already been doubted by Cliquennois (2005: 119), who hypothesized it might be related to the Madagscan tribe Achriopterini Günther, 1953 if the type-locality given as “ Madagascar ” was correct. Indeed, Parabactridium is most unlikely to belong in Cladoxerini , which is supported by certain characters mentioned in the original description by Redtenbacher (1908: 403) i.e. the small size (body-length including subgenital plate 90.0 mm), uni-spinose head, fairly short median segment, proportionally very short legs and abbreviated tarsi. Some of these characters rather suppose relation to the Madagascan subfamily Antongiliinae . Consequently, the genus is here only retained in Cladoxerini until its actual position is clarified. A confirmed placement of Parabactridium however remains as yet impossible.
A detailed diagnosis of the tribe would be premature, since investigation of supposedly closely related genera currently attributed to the Heteronemiinae : Paraleptyniini suggest Cladoxerini in its present recognition to be paraphyletic (→ see comments below). Hence, only a brief diagnosis mainly based on the type-genus Cladoxerus is provided here:
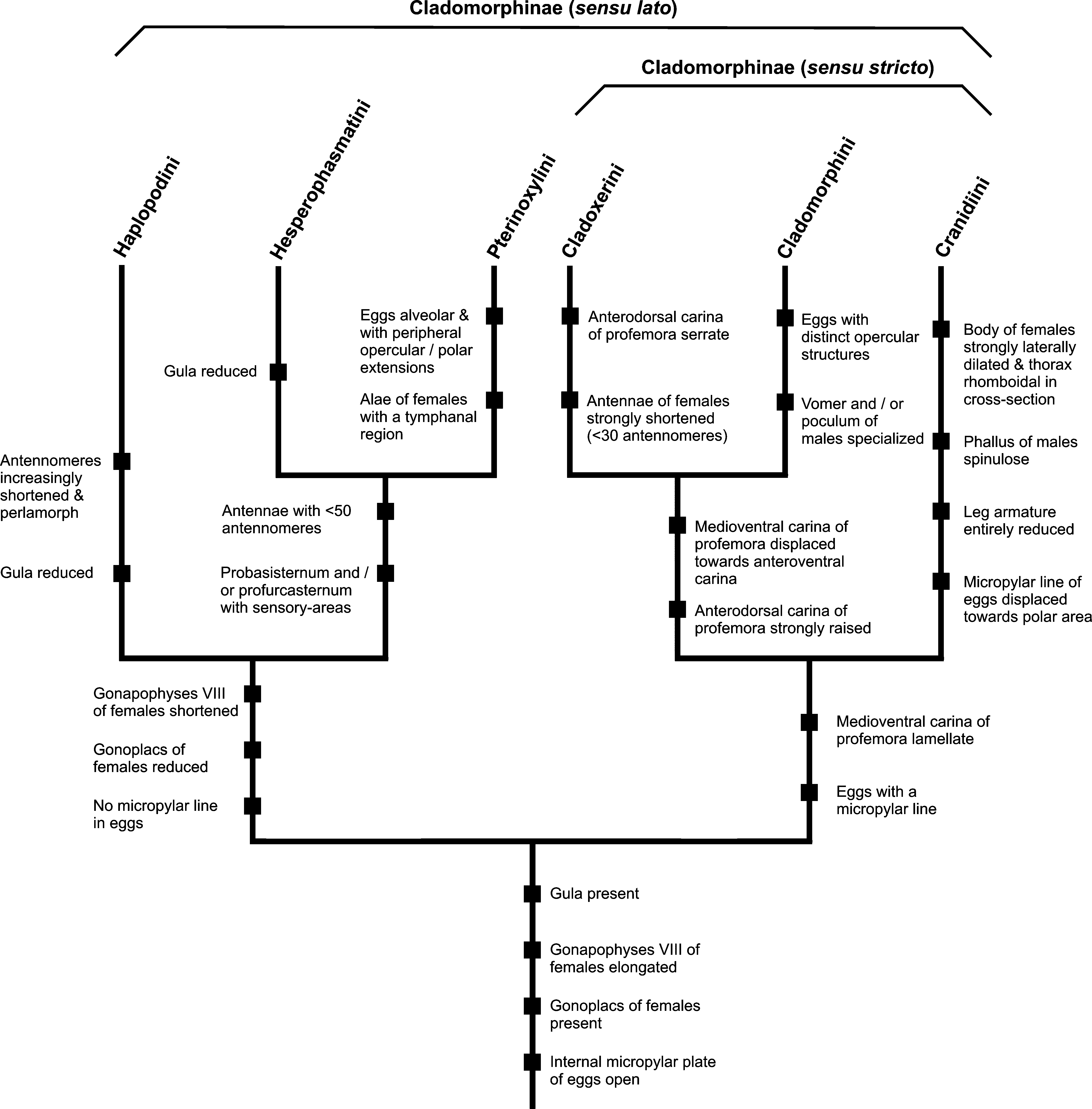
Members of Cladoxerini are well characterized and distinguished from all other five tribes currently attributed to Cladomorphinae by the dorsally serrate profemora of both sexes and shortened antennae, which consist of no more than 30 antennomeres. These are somewhat shorter than the profemora and have the antennomeres fairly elongate in ♂♂, while they are never considerably longer than the head and pronotum combined with the basal antennomeres strongly shortened and perlamorph in ♀♀ ( Fig. 3 View FIGURES 1 – 5 ). There are no ocelli. Both sexes are very slender and elongate stick-like insects, the ♀♀ being apterous, the ♂♂ (at least those of the type-genus Cladoxerus ) having scale-like tegmina and well developed alae ( Fig. 2 View FIGURES 1 – 5 ). The anal fan of the alae of ♂♂ is transparent. Most representatives are fairly large, ♀♀ of some species achieving body lengths of up to 240.0 mm including the long subgenital plate. The body surface is usually ± smooth and sub-glabrous. Occasionally there is a pair of spines on the head of ♀♀. A well developed and fairly large gula is present. Both sexes have a long median segment that is considerably longer than the metanotum. Females have a moderately developed praeopercular organ, which is formed by a median gap or slit near the posterior margin of abdominal sternum VII, and usually is marked by two bulge-like ridges or elevations. The subgenital plate of ♀♀ is strongly elongated, lanceolate, naviculate or spatulate and projects considerably over the apex of the abdomen ( Fig. 1 View FIGURES 1 – 5 ). The gonapophyses VIII are elongated, filiform but at best slightly exceed the posterior margin of the anal segment. Gonoplacs are present and moderately elongated. Males have the vomer well developed, sclerotized and elongately triangular with a pointed terminal hook. The poculum is small and angularly cup-shaped ( Fig. 4 View FIGURES 1 – 5 ). The profemora are distinctly triangular in cross-section, with the anterodorsal carina raised and ± distinctly serrate ( Fig. 1 View FIGURES 1 – 5 ); occasionally there may also be some teeth on the posteroventral carina. The medioventral carina is lamellate and strongly displaced towards the anteroventral carina. The mid and hind legs are trapezoidal in cross-section and ± prominently armed with a variable number of teeth or lobes. The tarsi are very elongate with the basitarsus carinate dorsally with the two dorsal carinae melted with another and considerably longer than the following tarsomeres; in ♀♀ the basitarsi often have a raised dorsal carina. Eggs are fairly small, longer than wide, laterally compressed, have the polar-area flattened and the anterodorsal portion of the capsule conspicuously convex. The capsule-surface is strongly sculptured. The micropylar plate is oval and fairly small, covering less than half the length of capsule. A median line is present. The operculum bears a raised but flattened structure, which is encircled by several conspicuous carinae on the outer surface ( Fig. 5 View FIGURES 1 – 5 ).
The shape of the profemora, having the lamellate medioventral carina strongly displaced towards the anteroventral carina, dorsally carinate basitarsi, and the well-developed gula of both sexes, the presence of gonoplacs and the moderately elongated, filiform gonapophyses VIII of ♀♀ as well as the presence of a median line in the eggs are shared with Cladomorphini , which supports close relation between these two tribes and suggests a sister-group relationship between Cladoxerini and Cladomorphini ( Fig. 409 View FIGURES 409 ). However, since close relation to the tribe Paraleptyniini Zompro, 2004 (currently in Heteronemiidae : Heteronemiinae ) is indicated by a good number of characters, Cladoxerini might by paraphyletic and any broader discussion on the true relationships of Cladoxerini would be premature at this point. The serrate anterodorsal carina of the profemora and increasingly shortened gonapophyses VIII of ♀♀ are likely to be autapomorphies of Cladoxerini . Whether the conspicuously shortened antennae of ♀♀ are plesiomorphic or apomorphic deserves further evaluation but is a priori interpreted as an autapomorphy of Cladoxerini in Fig. 409 View FIGURES 409 , since this character is unique within Cladomorphinae .
Distribution: Southeastern South America (SE-Brazil, Paraguay, Uruguay, Argentina & Bolivia).
Genera included:
1. Cladoxerus St. Fargeau & Audinet-Serville, 1828: 445. Type-species: Cladoxerus gracilis St. Fargeau & Audinet-Serville, 1828: 445, by monotypy. ( Figs. 1–2 View FIGURES 1 – 5 )
= Baculum Saussure, 1861: 127 . Type-species: Bacillus (Baculum) ramosum Saussure, 1861: 127 , by monotypy. [Originally described as a subgenus of Bacillus Latreille, 1825 ] n. syn.
= Abrachia Kirby, 1889: 503 . Type-species: Abrachia brevicornis Kirby, 1889 , by monotypy. [Synonymised with Baculum Saussure, 1861 by Zompro, 2004: 135]
= Ceratiscus Caudell, 1904: 188 . Type-species: Ceratiscus laticeps Caudell, 1904 , by original designation. [Synonymised with Baculum Saussure, 1861 by Zompro, 2004: 135]
2. Parabactridium Redtenbacher, 1908: 403 . Type-species: Parabactridium mirum Redtenbacher, 1908: 403 , by monotypy. [Presumably related to the Madagascan Antongiliini → see above]
3. Wattenwylia Toledo Piza, 1938: 4 . Type-species: Wattenwylia foliata Toledo-Piza, 1938: 6 , by original designation. [Here transferred from Pachymorphinae : Gratidiini ]

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |