
Halecium jaederholmi Vervoort, 1972
|
publication ID |
https://doi.org/10.11646/zootaxa.3790.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:BE6B199C-6E81-478A-8AC9-EB674B85FA35 |
|
DOI |
https://doi.org/10.5281/zenodo.4630889 |
|
persistent identifier |
https://treatment.plazi.org/id/3E6287E0-2959-FF98-2CA9-1CF73E8EFDB4 |
|
treatment provided by |
Plazi |
|
scientific name |
Halecium jaederholmi Vervoort, 1972 |
| status |
|
Halecium jaederholmi Vervoort, 1972 View in CoL
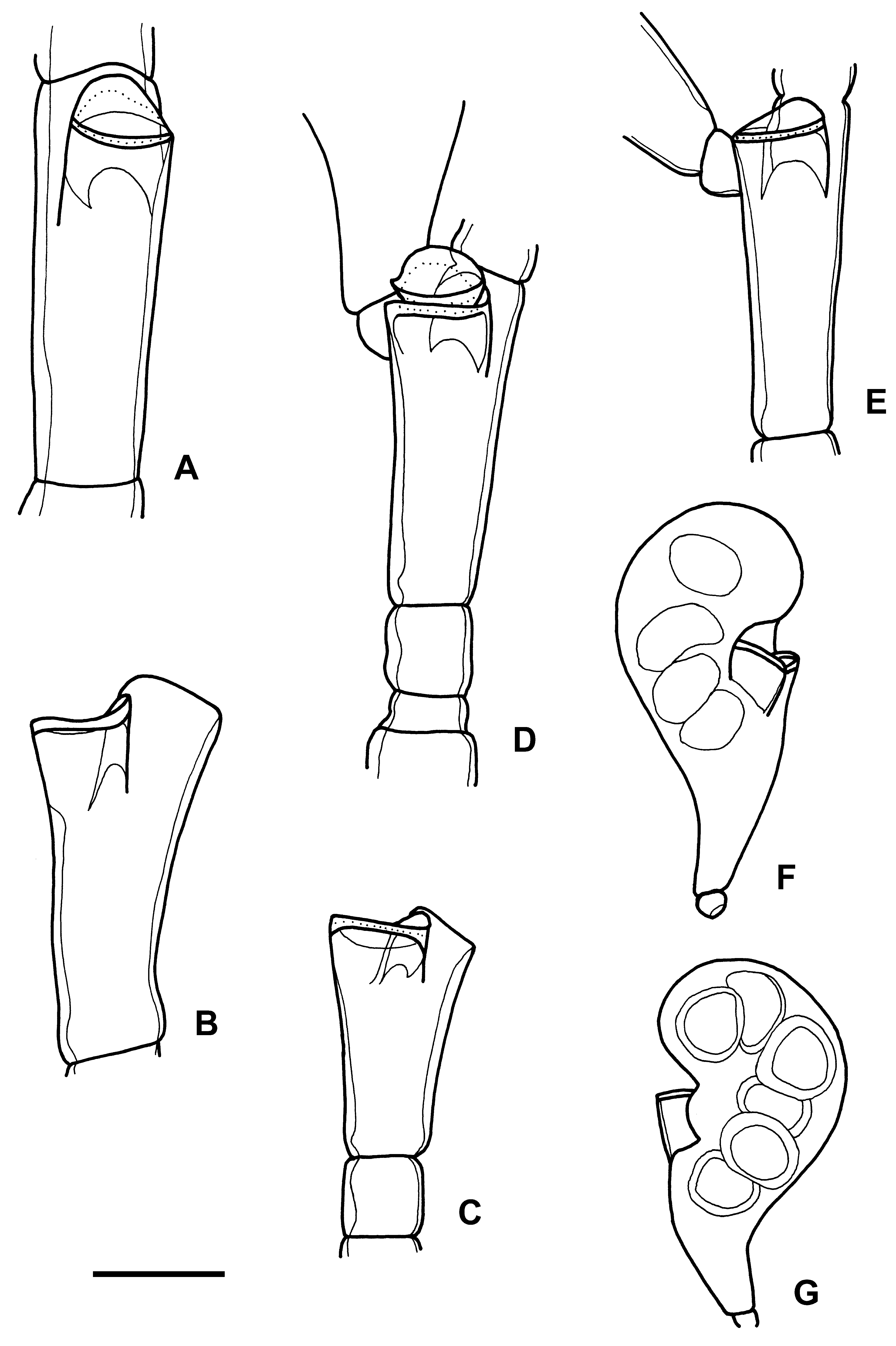
( Fig. 9 View FIGURE 9 )
Halecium jaederholmi Vervoort, 1972a: 21 View in CoL , fig. 2; 1972b: 343, fig. 2b, c; Millard, 1977: 10, fig. 3A–B; Stepanjants, 1979: 107, pl. 20 fig. 6; Branch & Williams, 1993: 12, fig.; Blanco, 1994a: 156; 1994b: 186–187; Vervoort & Watson, 2003: 86; Peña Cantero, 2004: 769; 2008: 455, fig. 1c, d; El Beshbeeshy & Jarms, 2011: 35 –38, fig. 4.
Halecium arboreum — Jäderholm, 1905: 11, pl. 5 fig. 4; Naumov & Stepanjants, 1962: 97; Stepanjants, 1972: 71.
Halecium robustum View in CoL — Ritchie, 1907: 524.
Halecium macrocephalum View in CoL — Ritchie, 1913: 18, fig. 4; Stechow, 1925: 402; Rees & Thursfield, 1965: 108.
Halecium View in CoL sp. Stepanjants, 1972: 72–73, fig. 17.
Material examined. Holotype, RMNH 7166, Stn Vema 17 RD 12, 44°19’S 59°52’W (shelf south-east of Peninsula Valdés, Argentina), 13–06–1961, 183– 366 m, several colony fragments up to 65 mm long.
Diagnosis. Strongly polysiphonic, dark-brown, irregularly branched stems, up to 160 mm long. Branches originating from hydrophore of primary hydrotheca. Hydrothecae alternately arranged in one plane. Hydrotheca at the end of short, adnate hydrophore. Hydrotheca barely widening distally; rim not everted. Adcauline hydrothecal wall adnate to internode and much larger than abcauline one. Hydrothecal aperture slightly directed downwards. A sessile secondary hydrotheca may be present. Female gonotheca kidney-shaped, with two hydrothecae, and up to six eggs. Cnidome consisting of microbasic mastigophores? and microbasic euryteles?
Description. The material belongs to at least six stems. Five stems arise from a shell fragment. Stems darkbrown, up to 5 mm in basal diameter, strongly polysiphonic all over the colony; only a few short side-branches monosiphonic. Polysiphony due to accessory tubes (sometimes in very high number).
Branching irregular, but alternate in one or two planes in some parts. Stems much branched, giving rise to lower-order branches, frequently strongly directed upwards (some completely vertical). Branches divided into internodes by slightly oblique alternate nodes ( Fig. 9 View FIGURE 9 A–E). Hydrothecae alternately arranged in more or less one plane. Adcauline hydrothecal wall almost reaching distal node of internode ( Fig. 9 View FIGURE 9 A–E).
Hydrothecae resting on adnate hydrophores ( Fig. 9 View FIGURE 9 A–E); ratio between adcauline length of hydrophore and diameter at diaphragm 0.5–1.0. Hydrotheca low, cylindrical, practically not widening distally ( Fig. 9 View FIGURE 9 A–E). Adcauline hydrothecal wall adnate and much larger than abcauline one, extending upwards on internode ( Fig. 9 View FIGURE 9 A–E). Hydrothecal aperture slightly directed downwards ( Fig. 9 View FIGURE 9 A–B,E). Sometimes a sessile secondary hydrotheca present ( Fig. 9 View FIGURE 9 D).
Female gonothecae present ( Fig. 9 View FIGURE 9 F–G), originating from lateral of hydrophore and resting on a short apophysis. Gonotheca kidney-shaped ( Fig. 9 View FIGURE 9 F–G), with two hydrothecae at the concave side. Up to six eggs observed ( Fig. 9 View FIGURE 9 G).
Measurements (in µm). Hydrothecae: diameter at aperture 180–210, diameter at diaphragm c. 190, height 15–25. Internodes: length 670–800, diameter just below distal node 150–250. Gonothecae: height 1370–1500, maximum diameter c. 700, basal pedicel c. 100 x 100. Cnidome: very abundant microbasic euryteles? with round ends [range 9–11 x 4–5, mean 10.0±0.5 x 4.5±0.3 (n=10); ratio, range 2.0–2.5, mean 2.2±0.1 (n=10)] and smaller microbasic mastigophores? with sharp ends (6–6.5 x 1.5).
Remarks. As indicated above Halecium brevithecum is similar to H. jaederholmi , and they could be conspecific. Both have polysiphonic colonies characterized by the extremely low hydrothecae. Both also have hydrothecae with the adcauline wall projecting upwards, being much larger than the rest, and adnate to internode. Additionally, the larger nematocysts seem to be of the same range.
There are, however, differences between both species. Although Watson (2008) indicated that “there is indication of desmocytes above the diaphragm” in H. brevithecum , I did not observe them. However, desmocytes are distinctly marked in H. jaederholmi . In addition, a sessile secondary hydrotheca is sometimes present in H. jaederholmi , whereas no secondary hydrotheca has been observed in H. brevithecum ; Watson (2008) also indicated “no marginal replications or linear series of hydrophores”. Halecium brevithecum is also characterized by the reddish stems, the distinctly longer first hydrothecate internode of the branches, and the more or less perpendicular arrangement of the lower-order branches in relation to the previous ones.
Millard (1977) undoubtedly found material of H. jaederholmi . She found female colonies agreeing perfectly with the type material. Colour from medium-brown to dark brown, with thick fascicled stems (up to 8 mm in diameter at base) up to 160 mm in height. Stems “branching irregularly and in all planes”. Millard indicated that “internodes and hydrophores have a structure exactly like that illustrated by Vervoort (1972a) except that no pseudodiaphragmata are present nor any secondary hydrophores”. As it has been shown above, there is actually no sign of pseudodiaphragm in the holotype. According to Millard (1977) up to six larvae are found in the gonothecae.
Branch & Willians (1993) also reported this species from the Marion and Prince Edwards islands area, but the few characters provided by them seem to be taken from Millard’s (1977) description, so it is not possible to be completely sure about the correctness of their identification.
Vervoort’s (1972b) material could correspond to another species. It consists of completely monosiphonic stems with yellowish perisarc. The hydrothecae seem to have the same height all around, and the adcauline side does not seem to fuse with the internode, contrary to what happens in the type material. Similarly, Stepanjants’s (1979: pl. 20 fig. 6) material, although polysiphonic in this case, may also correspond to a different species. Evidently, it would be necessary to re-examine all this material to assess whether it belongs to H. jaederholmi .
Due to the similarity between H. jaederholmi and H. brevithecum it is not possible to be completely sure about the proper identification of some previous records of H. jaederholmi , such as those by Jäderholm (1905), Ritchie (1907, 1913), Stepanjants (1979) and Peña Cantero (2008). Ritchie (1907: 524–525), for instance, indicated that in his material “the branches frequently have at their bases at least one athecate internode connecting the stem process with the hydrophore-bearing portions of the branch”. As indicated above, this is one of the few features that allow by now to characterize H. brevithecum . On the other hand, Ritchie (1907: 524) indicated that “rarely with a tier of one or two secondary hydrothecae” and that “inside of the limbus are situated small, light-refracting prominences”, whereas in H. brevithecum neither secondary hydrothecae nor desmocytes have been observed.
Ecology and distribution. It is difficult to establish its bathymetric and geographical distribution due to the uncertainty of some records. It has been found for sure at depths between 24 ( Vervoort 1972a) and 400 m ( Millard 1977). Gonothecae found in April ( Millard 1977) and June ( Vervoort 1972a).
Taking into account only the confirmed records, H. jaederholmi seems to have a sub-Antarctic distribution, being known from the Magellan area, in particular from the Atlantic entrance of the Magellan Strait and off Peninsula Valdés ( Vervoort 1972a) and from the Kerguélen area, between Possession and Cochons ( Millard 1977). Considering also the unconfirmed previous records, this species would have a Pan-Antarctic distribution, since it has been also reported from Antarctic waters, in particular off South Georgia ( Jäderholm 1905), Coats Land, in the Weddell Sea, and St Helena ( Ritchie 1907), Enderby Land ( Naumov & Stepanjants 1962), Arthur Harbor, Palmer Archipelago ( Vervoort 1972b), Davis Sea (Stepanjants 1972), the South Shetlands Islands ( Stepanjants 1979) and off Livingston and Deception islands ( Peña Cantero 2008). Additional sub-Antarctic records should be those by Jäderholm (1905), from the Falklands Islands area, Stepanjants (1979), from off Kerguélen and Crozet islands, Branch & Willians (1993), from the Marion and Prince Edwards islands area, and El Beshbeeshy & Jarms (2011), from the Patagonian shelf.
| RMNH |
National Museum of Natural History, Naturalis |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halecium jaederholmi Vervoort, 1972
| Peña Cantero, Álvaro L. 2014 |
Halecium jaederholmi
| El 2011: 35 |
| Pena 2004: 769 |
| Vervoort 2003: 86 |
| Blanco 1994: 156 |
| Branch 1993: 12 |
| Stepanjants 1979: 107 |
| Millard 1977: 10 |
| Vervoort 1972: 21 |
Halecium macrocephalum
| Rees 1965: 108 |
| Stechow 1925: 402 |
| Ritchie 1913: 18 |
Halecium robustum
| Ritchie 1907: 524 |
Halecium arboreum
| Naumov 1962: 97 |
| Jaderholm 1905: 11 |