
Prosaetes C. B. Wilson, 1907
|
publication ID |
https://doi.org/ 10.5281/zenodo.195669 |
|
DOI |
https://doi.org/10.5281/zenodo.6208850 |
|
persistent identifier |
https://treatment.plazi.org/id/404087F4-091C-FFF1-B0C1-DD1ECC16F841 |
|
treatment provided by |
Plazi |
|
scientific name |
Prosaetes C. B. Wilson, 1907 |
| status |
|
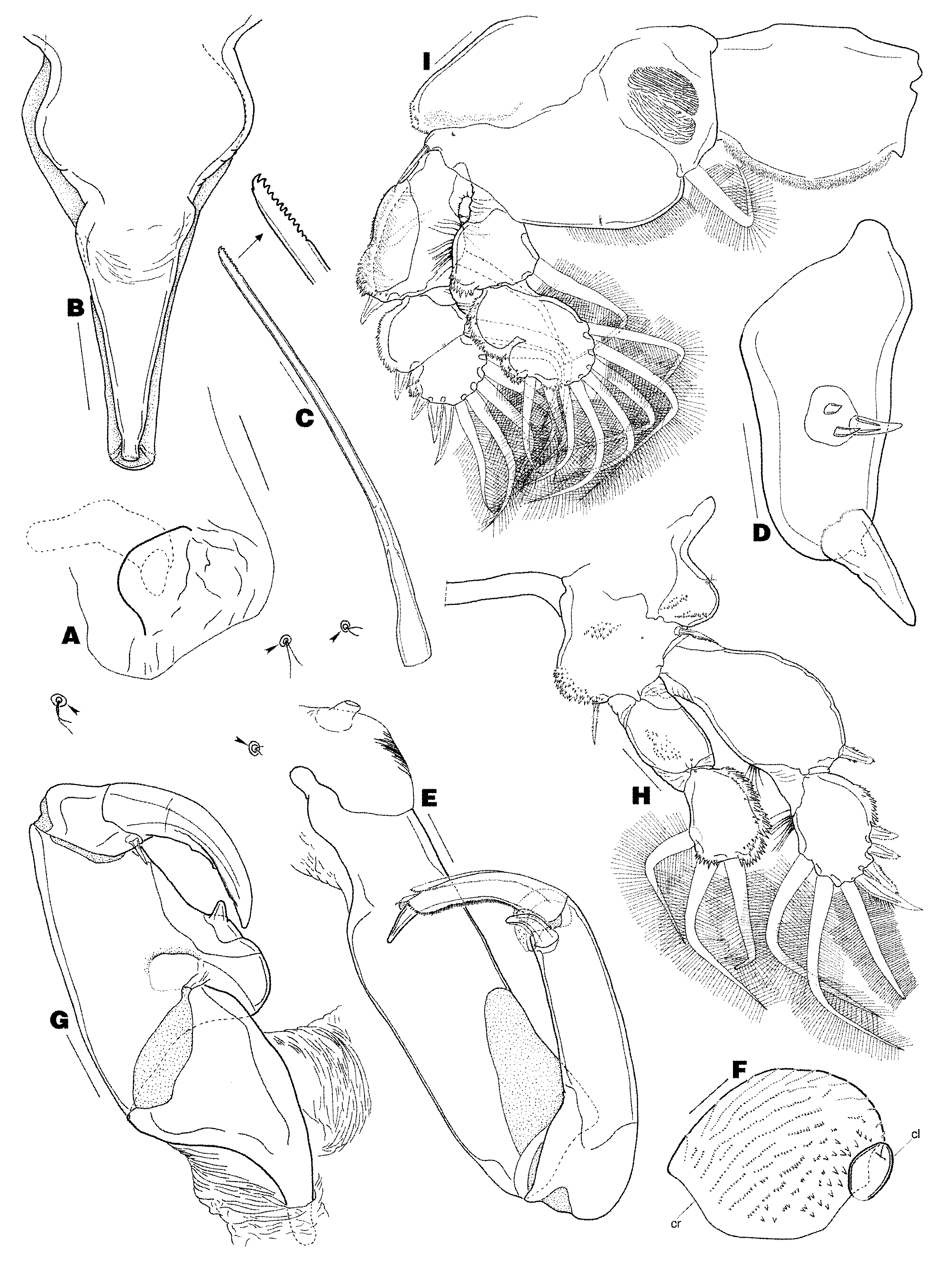
Amended diagnosis. Female body composed of caligiform cephalothorax incorporating first pedigerous somite only, first free thoracic somite comprising fused second and third pedigerous somites bearing welldeveloped lateral plate, second free thoracic somite consisting of fourth pedigerous somite bearing strongly serrated posterodorsal plate, suborbicular genital complex, and 1 free abdominal somite. Caudal ramus with 6 setae (seta I absent). Egg sacs uniseriate, straight. Paired frontal plates well-developed, without lunules. Antennule 2-segmented. Antenna 3-segmented, comprising coxa, basis, and 1-segmented endopod incorporating distal claw. Postantennal process vestigial. Oral cone elongate. Mandible modified into elongate stylet. Maxillule small, unilobate. Maxilla brachiform, 2-segmented, composed of syncoxa and basis; basis with crista and clavus in addition to calamus and canna. Maxilliped subchelate, comprising protopod and subchela consisting of free endopodal segment and claw. Leg 1 with 2-segmented rami. Legs 2 and 3 with 3- segmented exopod and 2-segmented endopod. Leg 4 rami incompletely 2-segmented. Leg 5 vestigial, unsegmented, situated on posteroventral surface of genital complex. Leg 6 rudimentary, represented by unarmed genital operculum. Male unknown. Parasite of the whale shark.
Type and only known species. Stasiotes rhinodontis Wright, 1876 by original designation.
Among the Cecropidae , Prosaetes shares 2 (vs. 3) free thoracic somites and relatively unmodified (vs. lamelliform) rami on legs 2–4 with Cecrops Leach, 1816 , Philorthagoriscus Horst, 1897 , and Luetkenia Claus, 1864 . Prosaetes can be easily distinguished from the these three genera, as well as Entepherus Bere, 1936 and Orthagoriscicola Poche, 1902 , by the absence of a posteromedian sinus on the genital complex (slight indentation in Cecrops ; highly indented in other genera), presence of a crista and clavus plus the usual calamus and canna on the maxillary basis ( Cecrops and Orthagoriscicola possess a crista plus calamus and canna; other genera with digitiform clavus plus calamus and canna), and retention of many plesiomorphic features such as cylindrical caudal rami (rounded in other genera except Philorthagoriscus ), 3 armature elements on the reduced maxillulary palp (1–2 in other genera except Philorthagoriscus ), well-developed plumose setae on legs 1–3 (leg setae greatly reduced in other genera), 3-segmented exopod on leg 2 (2- segmented in other genera), 8 setae on the distal endopodal segment of leg 2 (1 in Orthagoriscicola ; 7 in other genera), 3-segmented exopod on leg 3 (1-segmented in Orthagoriscicola ; 2-segmented in other genera), 6 setae on the distal endopodal segment of leg 3 (0 in Orthagoriscicola ; 5 in Entepherus ; 4 in other genera), incompletely 2-segmented rami in leg 4 (1-segmented in other genera), 9 elements on the distal exopodal segment of leg 4 (4 in Orthagoriscicola ; 8 in Philorthagoriscus ; 7 in other genera), and 5 elements on the distal endopodal segment of leg 4 (0 in Orthagoriscicola ; 3 in Cecrops and Philorthagoriscus ; 4 in other genera). A highly unusual feature exhibited by Prosaetes is the presence of 28 setae on the proximal antennulary segment, which diverges from the typical limit of 27 setae reported in the Cecropidae (see Benz & Deets 1988) and the majority of taxa in other caligiform groups (see Boxshall & Halsey 2004). This supernumerary state appears to be stable in Prosaetes as it was found in all dissected specimens examined in this study and has been reported, although infrequently, in other caligiform taxa [e.g., Caligus longipedis Bassett-Smith, 1898 and Synestius caliginus Steenstrup and Lütken, 1861 (see Ho & Lin 2004)].
Neotype material. A neotype ( USNM 1138870) was selected from Williams & Williams’ (1986) collection of 173 well-preserved, adult female specimens of Prosaetes rhinodontis deposited in the National Museum of Natural History ( USNM), Smithsonian Institution, Washington, D.C. Although Williams & Williams (1986) collected this material on 12 July, 1985 at the Okinawa Expo Memorial Park Aquarium (now named Okinawa Churaumi Aquarium), Japan, during a post-mortem examination of one R. typus specimen, historical records kept at this aquarium indicate that the host specimen was caught fortuitously on 10 July, 1985 in a net placed off the coast of Yonabaru-cho, southeastern Okinawa-jima Island (this whale shark was found dangling in the water with the caudal end entangled in the net near the surface and the opposite end bumping into the sea bottom), and then immediately transported alive, probably on a dump truck bed, to the aquarium. Based on this information we are certain that this whale shark specimen was infected with P. rhinodontis at the time of capture since it is highly unlikely that the large number of parasites (ca. 500–600 individuals), all at the adult stage, observed by Williams & Williams (1986) had accumulated on this host individual within a 3-day period. Further, the presence of a single pedunculate barnacle, Conchoderma virgatum (Spengler, 1790) , with a capitular length of 6.0 mm, attached to the dorsal surface of a P. rhinodontis individual collected by Williams & Willams (1986) indicates that this whale shark was infected by P. rhinodontis approximately 3 60 days prior to its death considering that the growth rate for C. virgatum has been estimated to range from 0.1 1.9 mm capitular length per day (see Eckert & Eckert 1987). The type locality of the neotype is accordingly designated the western North Pacific Ocean, off Yonabaru-cho (26°11΄N, 127°46΄E), Okinawa-jima Island, Okinawa Prefecture, Japan, rather than the Okinawa Churaumi Aquarium.
Neotype specimen is a non-ovigerous adult female preserved in 70% ethanol, light tan in color, and 6.02 mm long. Posterodorsal plate on fourth pedigerous somite with 6 large teeth on left side and 7 large teeth on right side. Genital complex with spermatophore cemented to each copulatory pore. Terminal endopodal segment of left leg 2 with abnormal innermost seta. All other features, such as the body shape and tagmosis, ornamentation of the cephalothorax and genital complex, and construction and armature of appendages, as described above.
Justification for neotype material. Edward P. Wright was a competent Irish naturalist, qualified in medicine, a lecturer in Zoology (1858–1868) at Trinity College, Dublin, Ireland, and Curator of the herbarium (1870 1910) at Trinity College ( Webb 1991). Prior to his latter appointment, Wright made a zoological expedition to the Seychelles in 1867, where he collected copepod samples from presumably two whale sharks which served as the basis for his description of Prosaetes rhinodontis . There is no indication that type specimens were deposited nor intended to be deposited in Wright’s original description of P. rhinodontis . Our recent correspondence with Mr. Mark Holmes, Keeper of Crustacea at the National Museum of Ireland, confirmed that specimens of P. rhinodontis are indeed absent from Wright’s crustacean collection housed at this museum.
A neotype is required for P. rhinodontis as confusion over the identity of this species has led to repeated misidentifications of this parasite species and the spurious establishment of a new species for this taxon. We have verified that all extant material, including the neotype, of P. rhinodontis presently available for study (more than 300 specimens) conform to Wright’s original description of this species. We anticipate that P. rhinodontis will be encountered more frequently in the immediate future due to the increasing number of whale sharks captured alive worldwide for display in public aquaria, high demand for whale shark products in Asia, and expansion of the whale shark ecotourism industry (see Colman 1997; Compagno 2001; Stevens 2007). The neotype and redescription provided herein of P. rhinodontis will, therefore, facilitate comparisons with future collections of this parasitic copepod from the whale shark.
Wright’s original material of P. rhinodontis was collected from presumably two whale sharks harpooned in waters off the Seychelles. The neotype was collected, on the other hand, from the same fish species captured in the western North Pacific Ocean, off the southeastern coast of Okinawa-jima Island, Japan. Kirtisinghe’s (1964) material of P. rhinodontis was collected considerably closer to the type locality, i.e. from a whale shark captured in the Indian Ocean off the coast of Colombo, Sri Lanka, but the whereabouts of his material remain unknown. Admittedly, selecting a neotype collected more than 8,000 km from the type locality is not ideal from a purist point of view. However, we believe it is warranted in this case since it is unlikely that new specimens of P. rhinodontis will be collected any time soon from the Seychelles, a country which recently afforded the whale shark the status of a protected species in Seychelles territorial waters under the auspices of “The Wild Animals (Whale Shark) Protection Regulations 2003” (see Rowat & Engelhardt 2007). In addition, we have established in this study that P. rhinodontis is a cosmopolitan and host specific parasite of the whale shark.
Biology. Some ectoparasitic copepod taxa are known to cluster together on their fish hosts in: a) singlefile [i.e. the cephalothorax of one copepod is positioned underneath the genital complex of an anterior copepod (e.g., see Kabata (1979) for clustering of Lepeophtheirus pectoralis (Müller, 1776) females on the inner surface of the pectoral fins of a pleuronectid flatfish)]; b) staggered formation [i.e. the lateral margins of the cephalothorax of one copepod are wedged underneath the lateral margins of the genital complexes of two anterior copepods (e.g., see Benz (1981) for aggregations of Pandarus satyrus Dana, 1852 individuals (primarily females) on the pectoral fins of Prionace glauca (Linnaeus) ]; or c) both single-file and staggered arrangements (e.g., see Dojiri & Cressey (1987) for clustering of Taeniacanthodes gracilis Wilson, 1935 individuals (mostly females) on the fins of Paralichthys squamilentus Jordan & Gilbert). Wilson (1907) and Kabata (1979) noted that large numbers of the cecropid Orthagoriscicola muricatus (Kr yer, 1837) tend to congregate on the skin of Mola mola (Linnaeus), but these authors did not mention the arrangement of individuals within the clusters. In this study we observed P. rhinodontis females aggregating together (arrowed in Figs 1B–D), albeit in an irregular pattern, mainly along the anterior margin of the sieve-like gill rakers of whale shark No. 31. Wright (1876), Kirtisinghe (1964), and Williams & Williams (1986) also found P. rhinodontis in the oral area of their respective whale shark specimen, but regrettably they did not provide details about the relative position of each copepod individual. Wilson’s (1944) record of this parasite from the skin of the whale shark requires verification. Benz (1981) argued that the aggregative behavior exhibited by P. satyrus may be biologically beneficial in terms of maximizing the limited area at the optimal attachment site, protecting copepod individuals from predation, assisting hydrodynamically in copepod attachment, and increasing the frequency of mating encounters between the small number of males and the comparatively more numerous females. Aggregation of P. rhinodontis females probably does not facilitate their attachment to the host since they typically clustered in an asymmetrical (Fig. 1E) rather than staggered or single-file arrangement as noted above for P. satyrus and L. pectoralis , respectively. Despite the lack of P. rhinodontis males among the present and historical collections, we speculate that P. rhinodontis females strategically aggregate to augment their reproductive fitness as evidenced by the large proportion (75% or 27/36) of ovigerous females among the in situ sample. Whether or not the absence of P. rhinodontis males among the samples obtained thus far is direct evidence that males occupy a microhabitat different from that of females, an indication that males preferentially mate with late juvenile or immature adult females and rapidly die thereafter, or a reflection of the rarity of collection of this gender remain to be determined.
Cecropids are generally considered to be relatively sedentary copepods as evidenced by the reduction and/ or loss of leg setae in both sexes, their inability to swim when placed in an aquarium, and the localized development of a shallow depression on the gill filaments, deep lesion on the skin, or proliferation of host tissue typically associated with prolonged attachment and feeding activities of these parasitic copepods ( Wilson 1907; Shiino 1965; Grabda 1973; Logan & Odense 1974; Kabata 1979, 1981; Benz & Deets 1988). In whale shark No. 31, the gill raker surface flanking P. rhinodontis individuals (Figs 1C–D) appears noticeably discolored, and in closer view slightly pinkish which most likely indicates epidermal tissue damage, as compared to the remaining gill raker area of this host individual and the monochromatic gill rakers of a whale shark free of P. rhinodontis (Fig. 1F). These observations suggest that P. rhinodontis not only grazes on the epithelium of the host’s gill rakers, probably by using its mandibles to rasp the surface of host tissue as is typical for other caligiform copepods ( Kabata 1974, 1982; Boxshall 2005), but also is, unlike other cecropids, relatively vagile. The latter is further substantiated by the fact that this parasite has well-developed plumose setae on legs 1–3 ( Figs 3 View FIGURE 3 H–I, 4A), presumably for facilitating swimming movements, and no gross host tissue responses (i.e., swelling, epithelial hyperplasia, or connective tissue encapsulation) were visible in the area immediately surrounding the parasite’s body (Fig. 1E).
Whale shark husbandry. Okinawa Churaumi Aquarium records show that whale shark No. 31 was placed with two other whale sharks in one (designated K-2) of three sea pens at the onset of its quarantine period on 8 November, 2007. The other two adjacent sea pens, K-1 and K-3, contained two whale sharks and one whale shark plus one giant manta ray (Manta birostris (Walbaum)), respectively. Whale shark No. 31 was subsequently transferred to K-1 on 11 December, 2007, and on 20 March, 2008 P. rhinodontis was detected for the first time in the oral region of this whale shark. This parasite was observed thereafter on whale shark No. 31 on 31 March, 12 April, 15 April, 29 April, 30 April, 2 May, and 9 May, 2008. The single whale shark (designated No. 16) held in K-3 died on 7 April, 2008; whale shark No. 31 was transferred to this sea pen on 12 April, 2008. On 8 May, 2008, one day before the death of whale shark No. 31, several P. rhinodontis individuals were observed for the first time in the oral area of two other captive whale sharks, one (designated No. 29) held in K-1 and the other (designated No. 30) in K-2. Prosaetes rhinodontis was not found on the giant manta ray held in K-3 at any time during the captivity period of whale shark No. 31. Based on this information and the fact that P. rhinodontis most likely has a direct life cycle typical of most copepods utilizing fish hosts (see Boxshall 2005), it appears that whale shark No. 31 was most likely infected with P. rhinodontis at time of capture and was accordingly the source of P. rhinodontis individuals that subsequently colonized whale shark No. 29 and No. 30. However, whale shark No. 29 and No. 30 being independently infected by P. rhinodontis at time of capture or transmission of P. rhinodontis from either whale shark No. 29 or No. 30, or both of these sharks, to whale shark No. 31 cannot be discounted.
The whale shark is a filter-feeder, employing both ram and suction filter-feeding behaviors to capture planktonic and nektonic prey, such as crab larvae, krill, copepods, sardines and anchovies, on its gill rakers ( Clark & Nelson 1997; Colman 1997; Compagno 2001; Taylor 2007). In association with this feeding habit, whale sharks in the wild have been observed to occasionally cough underwater, apparently as a mechanism to remove accumulated food particles from the gill rakers ( Colman 1997; Taylor 2007). Whale sharks held in captivity by the Okinawa Churaumi Aquarium exhibit a similar erratic coughing behavior (casual observations of M. Y.). Whale shark No. 31 in particular was observed coughing noticeably frequently, however, during periods of heavy infection by P. rhinodontis (casual observations of M. Y.), which may represent a sign of irritation, presumably in response to the parasite’s feeding and attachment activities on the gill rakers. As a caveat, we urge veterinary and husbandry staff overseeing whale sharks at public aquaria to regularly examine the oral cavity of these animals for P. rhinodontis during and after a lengthy quarantine period to prevent the inadvertent introduction of this parasite into display aquaria, particularly considering that this parasite can survive in an aquarium setting (see Williams & Williams 1986), can persist on captive whale sharks for a relatively lengthy period, has the ability to proliferate amongst captive whale sharks, has the potential to cause undue stress to captive whale sharks, and in view of its infection site, may reduce the feeding efficiency of a heavily infected whale shark. Further, treatment protocols to control or eradicate this parasite have not yet been developed.
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |