
Hyperolius cinereus Monrad, 1937
|
publication ID |
https://doi.org/10.11646/zootaxa.3635.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:E9955681-99A8-4618-A4CC-DD838F4294CE |
|
DOI |
https://doi.org/10.5281/zenodo.6146334 |
|
persistent identifier |
https://treatment.plazi.org/id/421187CC-FFBC-FF9B-FF21-2316FAD2E08D |
|
treatment provided by |
Plazi |
|
scientific name |
Hyperolius cinereus Monrad, 1937 |
| status |
|
Hyperolius cinereus Monrad, 1937 View in CoL
Ashy reed frog
Figs 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6
Material examined. See Appendix A.
Holotype. MHNC 90.0857, a male holotype from “Kalukembe” (= Calaqumbe), Angola (examined from photographs only).
Comment. Two specimens were available to Monard (1937). However, he specifically stated that he provisionally refers the specimen from “Bimbi” (= Bimbe) to the species: "… un jeune du Bimbi, différant en plusieurs points, appartient peut-être aussi à cette espèce; … a young specimen from Bimbi, different in several points, may also belong to this species". We therefore do not consider it part of the type material, although we assign it provisionally to the same species.
Diagnosis. A species referred to Hyperolius due to the following characteristics (Schiøtz 1999, Channing 2001): pupil horizontal; vocal sac present in male, with the gular flap oval with lateral and posterior margins free; terminal discs on fingers and toes expanded, rounded; tympanum hidden; skin smooth; and molecular monophyly for the 16S mitochondrial marker with other Hyperolius taxa.
Hyperolius cinereus can be distinguished from other Hyperolius in Angola and adjacent regions by: ventrum uniform yellow without darker pigmentation (heavily pigmented in H. steindachneri ); throat without spines (spinose in H. spinigularis males); no pale triangle on snout (present in H. ocellatus ); no light heel spot (present in H. mitchelli ); no translucent green belly skin (present in H. nasutus complex); no pale canthus and dorsolateral lines (red in H. kachalolae , dark in H. kivuensis , H. vilhenai , and H. bocagei ); back uniform without patterns or bands (distinct from quite variable patterns in H. chelaensis , H. major , H. pictus , H. pollii , H. seabrai , H. punctulatus and the H. angolensis complex; at least two stripes in H. quinquevittatus ; darker anterior coloration in H. bicolor ; brown with lateral and dorsal stripes in H. gularis ); no light lateral band or scattered dorsal spots (present in H. cinnamomeoventris ).
In the phylogenetic analysis (see Results) H. cinereus is sister to H. chelaensis , from which it shows an uncorrected p-distance of 3.7%. It further differs from H. chelaensis in having: a brightly coloured (uniform green to bluish) dorsum; bright red inner thighs; large yellow protruding eyes; having no dorsolateral stripes; smaller average size (male 21.0 mm vs 24.6 mm; female 24.6 vs 31.5 mm); in vocalisation having a higher average frequency advertisement call (3300 hz vs 2500 hz), a shorter initial creak call (160 ms vs 500 ms), followed by a higher burst of short chirp calls (2–14 vs 4–5); and finally it differs in habitat preference (grassland vs Afromontane forest stream).
Redescription of male. Based on an adult male PEM A9216 (measurements in mm): habitus squat, widest at midbody, tapering to the groin; head comparatively small (HL/SUL 0.3, HW/SUL 0.3), not wider than trunk, slightly wider than long (HW/HL 0.9); snout long ( END +NSD/HL 0.5), blunt in dorsal view, round in profile, slightly projecting beyond lower jaw, wider than long; canthus rostralis distinct, rounded; loreal region almost vertical, slightly concave; nostril directed dorsolaterally, situated much closer to tip of snout than to eye ( END / NSD 1.5), separated from each other by distance not much greater than distance between eye and nostril ( IND / END 1.1); eyes directed anterolaterally, protruding, relatively small (ED/HL 0.4); eye diameter shorter than snout (ED/ END +NSD 0.8); interorbital distance greater than internarial distance (IOD/ IND 1.2); tympanum not visible externally; tongue long 4.4, and narrow (2.1 at widest point), free for approximately 60% of length, forked distally for approximately 20% of length; vocal sac single; gular flap consisting of two medially arranged, subcircular areas of thickened skin, immediately adjacent to each other, anterior part cream-coloured, larger, more granular, and thicker than posterior white-coloured part; in resting position only anterior part visible from ventral; dorsal surfaces of head, trunk and limbs are smooth; ventral surface of limbs and gular smooth, chin and abdomen slightly more granulated; supratympanic fold absent.
Forelimbs slender; hand moderately large (HND/SUL 0.2); tips of fingers enlarged into broad oval disks, each with circummarginal groove; relative length of fingers: I<II<IV<III; longest finger 4.8; subarticular tubercles rounded, well developed, with one on fingers I and II, two on fingers III and IV, with proximal tubercle on finger IV hardly discernible; webbing between fingers minimal, no inner or outer metacarpal tubercles present; palmar tubercles absent; fingers without subdigital tubercles; nuptial pads or asperities absent. Hind limbs slender, moderately long (LEG/SUL 1.6); tibio-tarsal articulation reaching to posterior level of the eye when legs are adpressed to body; relative length of toes: I<II<III<V<IV; longest toe 7.7; discs of toes smaller than those of fingers; subarticular tubercles: one on toes I and II, two on toes III and V, and three on toe IV; foot webbing formula: I (0.25), II i/e (1-0.25), III i/e (1-0.25), IV (1), V (0.25); inner metatarsal tubercle small and indistinct; outer metatarsal tubercle absent.
Dorsum uniform lime green/yellow without any markings; ventrum, legs and feet uniform yellow without any markings; inner thighs red, tip of toes and fingers yellow; distinct separation between ventral and dorsal colouration (thus bicoloured); iris bright yellow. In preservative the yellow ventrum fades to light beige, and the dorsum becomes ashy-blue.
Further measurements: SUL 21.7; HL 6.7; HW 5.9; ED 2.5; END 1.9; NSD 1.3; IOD 2.3; IND 2.0; THL 10.3; TL 10.5; FL 14.3; HND 4.8; LEG 35.1.
Description of female. Female previously unknown, based on PEM A9219 ( Fig. 4 View FIGURE 4 B; measurements in mm); the female resembles the male but differs in: larger size; nostril closer to snout ( END /NSD 2.5); palm of hand more granular. Female more brightly coloured than males, dorsum blue/green, with light white band separating the dorsum and ventrum; ventrum bright yellow, inner thighs infused with bright red, inner arms and feet orange, upper arms and legs same colouration as dorsum, discs on toes and fingers orange, iris yellow. Sexual dimorphism follows that of other Hyperolius species, e.g. absence of vocal sac and gular flap in females.
Further measurements of female: SUL 28.7; HL 8.9; HW 8.4; ED 3.2; END 2.7; NSD 1.1; IOD 2.8; IND 2.5; THL 12.0; TL 12.2; FL 19.1; HND 7.2; LEG 43.3.
Variation. Maximum male SUL 26.0 (median: 21.0) and maximum female SUL 28.7 (median: 24.6). Further body dimensions and ratios are summarised in Table 3 View TABLE 3 . All males and females have similar colouration to that described above, except for one male having olive green dorsum.
Tadpole. Previously unknown (Channing et al. 2012). Description based on single specimen from tadpole lot PEM T558 (measurements in mm) at Gosner stage 38. Sequence divergence between adult and tadpole material varies from 0.0 to 0.4% indicating that tadpoles were conspecific with the adults. Lateral view ( Fig. 5 View FIGURE 5 A): Body round to ovoid; body little wider than high (BH/BW 0.9); snout oblique; mouth directed near ventral; nasals rounded, very small, positioned laterally, closer to snout than to eyes (SND/NOD 0.8); eyes moderately sized (ED 12% of BL), positioned and directed laterally; spiracle sinistral, tubular, small, joined to body wall, positioned laterally and situated closer to vent than snout (SS 58% of BL), oval opening directed slightly upwards, at the height of the middle of the lower part of the caudal muscle; tail just over twice the body length (TAL/BL 2.1) and two thirds of total length (TAL/tTL 0.7); tail musculature moderate (TMH 67% of BH and 62% of MTH), tapering gradually to a slightly pointed tail tip; tail fins of moderate size, deepest midway along tail; upper fin not extending onto body, slightly convex to the end of the tail; lower fin slightly convex to the end of tail. Dorsal view ( Fig. 5 View FIGURE 5 D): Body oval, little wider than high (BH/BW 0.9), widest just behind the spiracle opening; snout rounded; nasals widely spaced; interorbital distance nearly equal to internarial distance (IOD/ IND 1.10); tail muscle width 40% of body width (TMW/BW 0.4). Ventral view ( Fig. 5 View FIGURE 5 C): Eyes bulging and not visible in ventral view; vent positioned supramarginal, dextral and short with an oval opening. Oral disc ( Fig. 5 View FIGURE 5 B): Positioned and directed near ventral, moderately small (ODW 24% of BW); LTRF: 1/3(1), third posterior row 75% the length of the first row; jaw sheets moderately pigmented, capsulated and rounded; lateral process moderately short; single row of posterior marginal papillae, and a single row of anterior marginal papillae; dorsal gap free of papillae (up to 75% free); posterior corners with a few submarginal papillae; lateral margins of oral disc slightly indented. Colouration pattern: Body dark brown-yellow with randomly scattered dark spots; dorsum dark; ventrum paler with scattered darker pigmentation; internal collided intestine not visible through skin; tail musculature more heavily pigmented than fins; three distinct darker pigmented stripes running from anterior part of tail muscle to about midway of tail; fins mottled and translucent. Variation among tadpoles include tail tip being darker pigmented, intestinal coils visible through ventral skin and the most posterior labial tooth rows having variable lengths. Measurements: tTL 38.0; BL 12.2; TAL 25.8; BH 6.5; BW 7.6; TMW 3.0; MTH 7.1; TMH 4.4; IOD 3.5; IND 3.2; NOD 2.0; SND 1.6; SOD 3.6; SS 7.1; ED 1.5; ODW 1.8.
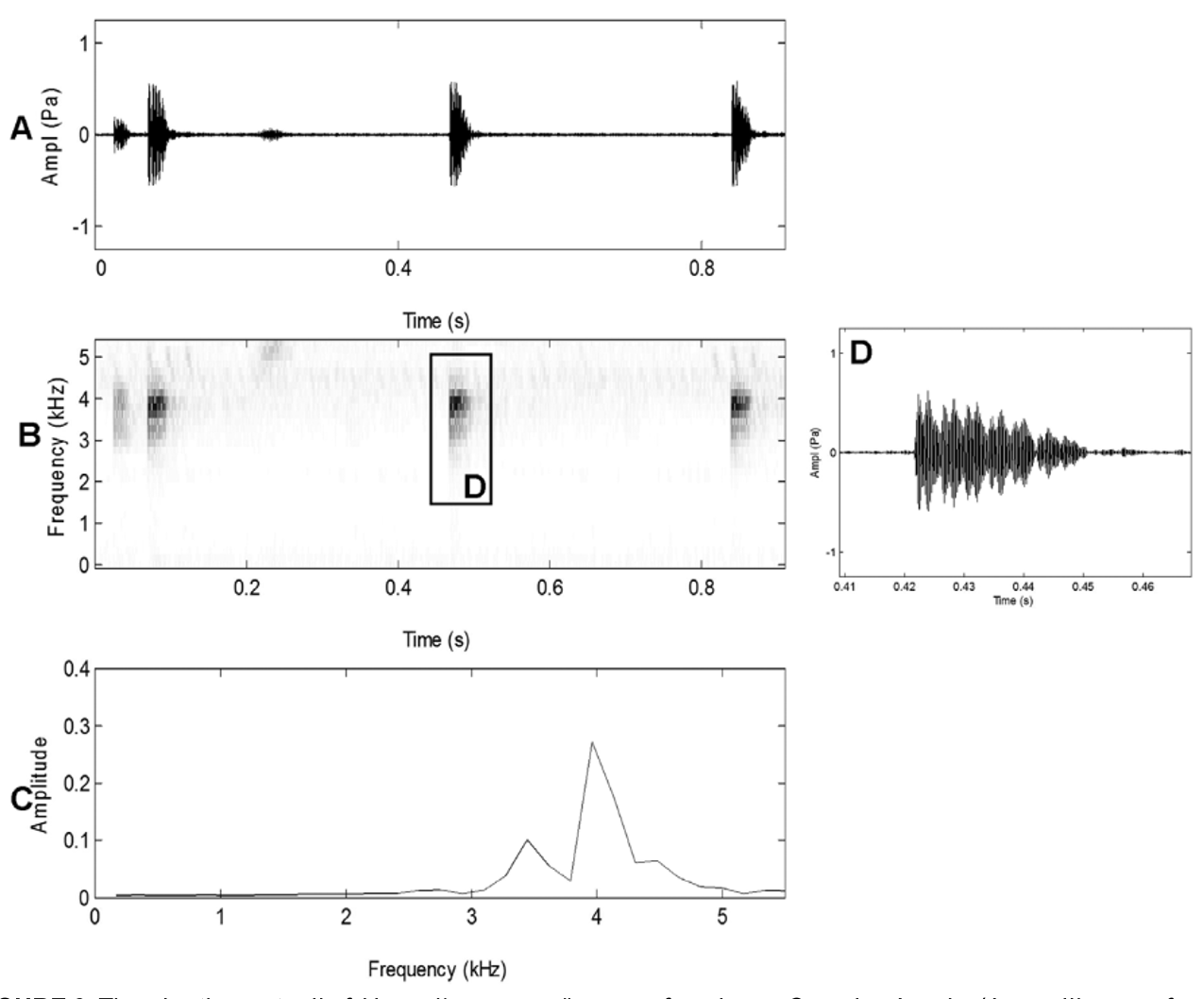
Advertisement call. Air temperature at time of recording 12–16 °C. The call (n = 22) usually comprises a series of two to fourteen notes emitted at a rate of 3.4 ± 1.0 (2.2–5.0) per second (mean ± SD [range], Fig. 6 View FIGURE 6 A). Usually, the series consists of a single initial creak with a pulse group composed of 4–8 (median: 6) pulses, and a length of 159.6 ± 92.5 (51–355) ms, followed by a series of 2–14 (median: 5) chirps with a pulse group composed of 2–4 (median: 3) pulses, and a length of 26.6 ± 10.7 (10–73) ms ( Fig. 6 View FIGURE 6 B–E). Dominant frequency ranged between 2842 and 3704 Hz ( Fig. 6 View FIGURE 6 C).
Natural history. Specimens were collected whilst calling at night, or basking on vegetation during the day. Found in grassland ‘dambo’, with water levels about 50cm deep ( Fig. 4 View FIGURE 4 E). At Humpata, specimens were found near a small flowing stream in montane grassland. Males were calling 30–50 cm from the water surface on grass stems. The following frog species were found sympatric with this species: Hyperolius angolensis , H. benguellensis , Amietia angolensis and Phrynobatrachus mababiensis .
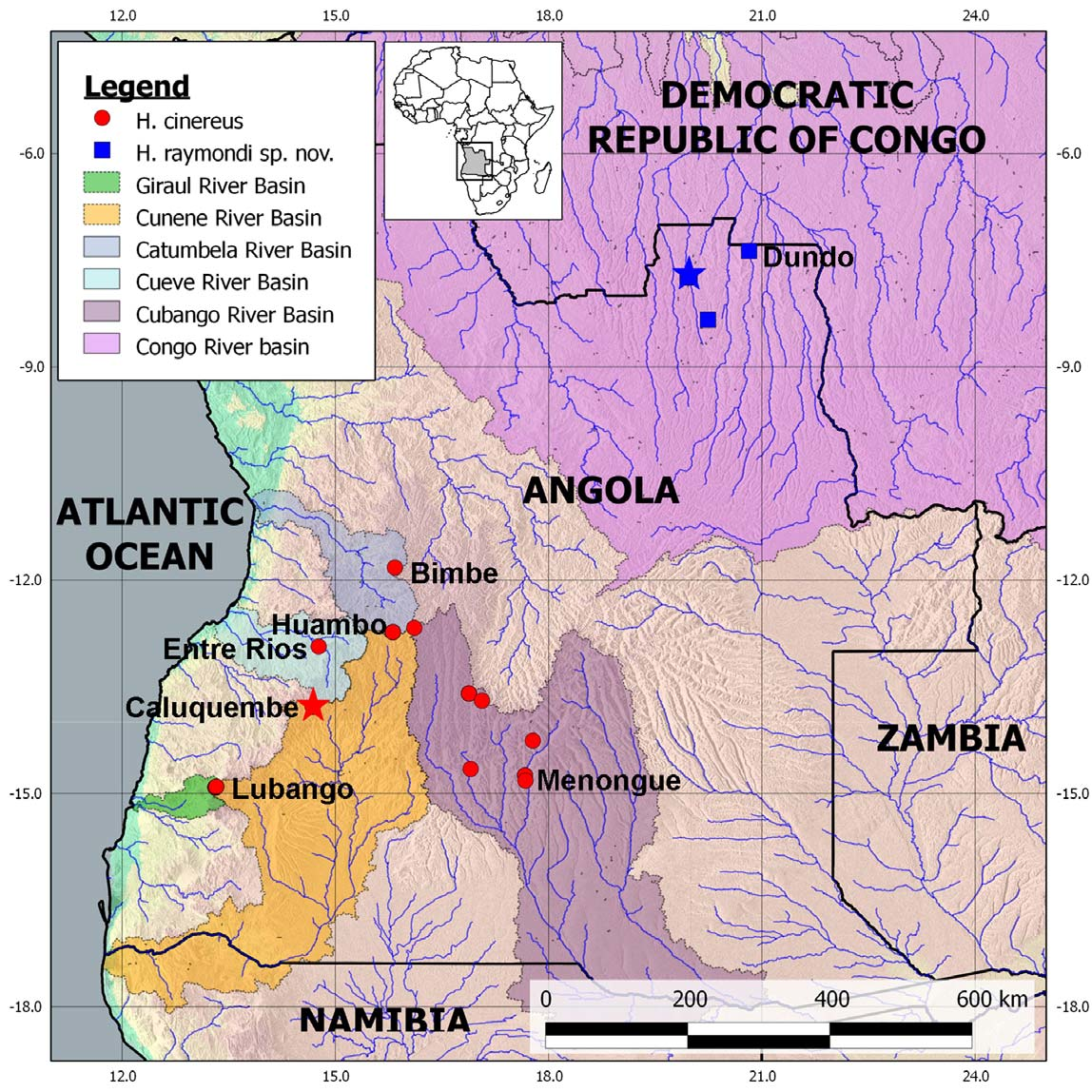
Distribution. Historically only known from three localities in south-central Angola (e.g. Caluquembe, Bimbe and Entre Rios). We report here an additional eight localities, extending the distribution of the species 200 km south and 350 km east of the type locality ( Fig. 1 View FIGURE 1 ). It is therefore more widespread than previously known. The species is restricted to high laying areas of Angola (above 1200 m above sea level).
Conservation status. This species is endemic to Angola. The current IUCN status of this species is Data Deficient. As the species is known to have a wide distribution (extent of occurrence is ~90 0 0 0 km2, area of occupancy is ~6 800 km 2), is known from more than 10 localities (quarter degree grid squares), and there is no evidence of population declines or fluctuations, the species should be considered as Least Concern (IUCN 2001). It appears to adapt well to habitat disturbance as it was found in flooded agricultural fields near Huambo.
Hyperolius cinereus— Laurent 1964 (part), Schiøtz 1999 (part), Channing 2001 (part).
Holotype. An adult male (PEM A10049, Fig. 7 View FIGURE 7 A) deposited in the Port Elizabeth Museum (PEM), South Africa. Collected 30 April 2012, by W. Conradie, W.R. Branch, P. Vaz Pinto, S. Baptista & N. Baptista from a dambo near the expedition base camp, Lagoa Carumbo, Lunda Norte Province, Angola ( 7°44’39.2” S; 19°57’16.8” E, 0719DB, 813 m above sea level).
Allotype. An adult female (PEM A10066, Fig. 7 View FIGURE 7 B), identical collecting details as Holotype.
Paratypes. 16 males (PEM A10040–10048, A10050 View Materials –10056), identical collecting details as Holotype.
Additional material. PEM A10057 [ 1 male] collected on 5 May 2010, by P. Vaz Pinto, S. Baptista & N. Baptista from the headwaters of Lovua, north of village Capaia, Lunda Norte Province, Angola ( 8°20’18.5” S; 20°14’33” E, 0820AC, 154 m above sea level). PEM T517 [2 tadpoles] collected on 2 May 2012, by W. Conradie & W.R. Branch from the margins Lagoa Samokaza east of Lagoa Carumbo, Lunda Norte Province, Angola ( 7°44’18.0” S; 19°59’13.0” E, 0719DB, 867 m above sea level).
Diagnosis. A species referred to Hyperolius due to the following characteristics (Schiøtz 1999, Channing 2001): pupil horizontal; vocal sac present in male, with the gular flap oval and lateral and posterior margins free; terminal discs on fingers and toes expanded, rounded; tympanum hidden; skin smooth; and molecular monophyly for the 16S mitochondrial marker with other Hyperolius taxa.
Hyperolius raymondi sp. nov. can be distinguished from other Hyperolius in Angola and adjacent regions by: ventrum uniform yellow without darker pigmentation (heavily pigmented in H. steindachneri ); throat without spines (spinose in H. spinigularis ); no pale triangle on snout (present in H. ocellatus ); no light heel spot (present in H. mitchelli ); no translucent green belly skin (present in H. nasutus complex); two dorsolateral stripes present (distinct from quite variable patterns in H. chelaensis , H. major , H. pictus , H. pollii , H. seabrai , H. punctulatus and the H. angolensis complex); uniform green colouration with darker bands present (bands absent in H. cinereus , darker anterior coloration in H. bicolor , brown with lateral and dorsal stripes in H. gularis ); dorsolateral stripes darker than background (distinct from H. quinquevittatus and H. cinnamomeoventris ); no light stripe below dark dorsolateral band ( H. kivuensis ).
In the phylogenetic analysis (see Results) it is sister to H. cinereus Monrad 1937 , from which it differs by 3.7% sequence divergence. It further differs from H. cinereus in having: up to two dark dorsolateral lines in males (vs no dorsal markings); a longer snout; faster call rate (4.4 vs 3.4 average notes per second); higher average frequency advertisement call (3900 hz vs 3300 hz); no initial creak call; lower number burst of short chirp calls (2–3 vs 2–14); lower number of pulse groups (1 vs 3); and tadpoles having two posterior rows of papillae (vs one row).
Description of Holotype. (measurements in mm) Habitus squat, widest at midbody, tapering to the groin; head comparatively small (HL/SUL 0.3, HW/SUL 0.3), not wider than trunk, slightly wider than long (HW/HL 0.9); snout moderately long ( END +NSD/HL 0.5), blunt in dorsal view, round in profile, slightly projecting beyond lower jaw, wider than long; canthus rostralis distinct, rounded; loreal region almost vertical, slightly concave; nostril directed dorsolaterally; situated closer to tip of snout than to eye ( END /NSD 1.2), separated from each other by distance not much greater than distance between eye and nostril ( IND / END 1.1); eyes directed anterolaterally, protruding, relatively small (ED/HL 0.3); eye diameter shorter than snout (ED/ END +NSD 0.7); interorbital distance greater than internarial distance (IOD/ IND 1.3); tympanum not visible externally; tongue long 5.5, and narrow (2.7 at widest point), free for approximately 55% of length, forked distally for approximately 25% of length; vocal sac single; gular flap consisting of two medially arranged, subcircular areas of thickened skin, immediately adjacent to each other, anterior part cream-coloured, larger, more granular, and thicker than posterior white-coloured part; in resting position only anterior part visible from ventral; dorsal surfaces of head, trunk and limbs are smooth; ventral surface of limbs and gular smooth, chin and abdomen slightly more granulated; supratympanic fold absent.
Forelimbs slender; hand moderately large (HND/SUL 0.2); tips of fingers enlarged into broad oval disks, each with circummarginal groove; relative length of fingers: I<II<IV<III; longest finger 4.9, subarticular tubercles rounded, well developed, with one on fingers I and II, two on fingers III and IV, with proximal tubercle on finger IV hardly discernible; webbing between fingers minimal, no inner or outer metacarpal tubercles present; palmar tubercles absent; fingers without subdigital tubercles; nuptial pads or asperities absent. Hind limbs slender, moderately long (LEG/SUL 1.6); tibio-tarsal articulation reaching to posterior level of the eye when legs are adpressed to body; relative length of toes: I<II<III<V<IV; longest toe 7.9; discs of toes smaller than those of fingers; subarticular tubercles: one on toes I and II, two on toes III and V, and three on toe IV; foot webbing formula: I (0.25), II i/e (1-0.25), III i/e (1-0.25), IV (1), V (0.25); inner metatarsal tubercle small and clear; outer metatarsal tubercle absent.
Dorsum uniform lime green with two distinct dark dorsolateral stripes on each side, lower dorsolateral stripe extending from tip of snout through the eye backwards to the groin; the upper dorsolateral stripe usually originates at the anterior corner of the upper eyelid, extending posterior and almost merging with the lower stripe; black spots of variable size occur on the posterior dorsum between the upper dorsolateral stripes; scattered small asperities on head; ventrum, and ventral surfaces of limbs uniform yellow, without markings; inner thighs red; tip of toes and fingers yellow; no distinct separation between ventral and dorsal colouration; upper jaw marked with broken black markings; upper thighs and forearms with scattered black markings on lime green background; palm of hands and soles of feet scattered with numerous small black asperities; iris bright yellow. In preservative the yellow colour of ventrum fades to light beige and the dorsum becomes ashy-blue with paired black dorsolateral stripes.
Further measurements: SUL 21.2; HL 6.8; HW 6.0; ED 2.5; END 1.8; NSD 1.6; IOD 2.6; IND 2.0; THL 10.3; TL 9.6; FL 13.9; HND 4.9; LEG 33.8.
Variation. Maximum male SUL 22.6 (median: 19.8) and female SUL 22.7 (only one specimen). Laurent (1964) reports the maximum male as 23.3 and female as 28.1. Further body dimensions and ratios are summarised in Table 3 View TABLE 3 . All male paratypes possess similar dorsal and ventral colouration to the holotype, but with individual variation in dorsolateral stripes ( Fig. 7 View FIGURE 7 ). The only female ( paratype PEM A10066; Fig. 7 View FIGURE 7 B) is indistinguishable in colouration from female H. cinereus , with a uniform blue-green dorsum, yellow ventrum and red inner thighs. Two juveniles collected were also bicolour , with darker lateral markings restricted to the snout.
Tadpole. Based on single specimen from tadpole lot PEM T517 (measurements in mm) at Gosner stage 36. Sequence divergence between adult and tadpole material varies from 0.2 to 0.6%, indicating that tadpoles were conspecific with the adults. Lateral view ( Fig. 8 View FIGURE 8 A): Body round to ovoid; body little wider than high (BH/BW 0.8); snout oblique; mouth directed near ventral; nasals rounded, very small, positioned laterally, closer to snout than to eyes (SND/NOD 0.8); eyes moderately sized (ED 16% of BL), positioned and directed near lateral; spiracle sinistral, tubular, small, joined to body wall, positioned laterally and situated closer to vent than snout (SS 69% of BL), oval opening directed slightly upwards, at the height of the middle of the lower part of the caudal muscle; tail is just over twice the body length (TAL/BL 2.2) and two thirds of total length (TAL/tTL 0.7), tail musculature broad (TMH 90% of BH and 86% of MTH), tapering gradually to a slightly pointed tail tip; tail fins of moderate size, deepest midway down tail; upper fin not extending onto body, slightly convex to the end of the tail; lower fin slightly convex to the end of tail. Dorsal view ( Fig. 8 View FIGURE 8 D): Body oval, little wider than high (BW/BH 1.3), widest just behind the spiracle opening; snout rounded, slightly pointed; nasals widely spaced; interorbital distance nearly equal to internarial distance (IOD/ IND 1.0); tail muscle width 48% of body width (TMW/BW 0.5). Ventral view ( Fig. 8 View FIGURE 8 C): Eyes bulging and not visible in ventral view; vent positioned supramarginal, dextral and short with an oval opening. Oral disc ( Fig. 8 View FIGURE 8 B): Positioned and directed near ventral, moderately small (ODW 29% of BW); LTRF: 1/3(1), third posterior row 75% the length of the first row; jaw sheets moderately pigmented, capsulated and rounded; lateral process moderately short; double row of posterior marginal papillae, and a single row of anterior marginal papillae; dorsal gap free of papillae (up to 75% free); posterior corners with a few submarginal papillae; lateral margins of oral disc slightly indented. Colouration pattern: Body dark brown with randomly scattered dark spots; dorsum dark; ventrum paler with scattered darker pigmentation; internal collided intestine slightly visible through skin; tail musculature more heavily pigmented than fins; three distinct darker pigmented stripes running from anterior part of tail muscle to approximately midway in tail; fins mottled and translucent. Measurements: tTL 35.0; BL 10.9; TAL 24.1; BH 4.8; BW 6.3; TMW 3.0; MTH 5.0; TMH 4.3; IOD 3.2; IND 3.1; NOD 2.0; SND 1.6; SOD 3.5; SS 7.5; ED 1.7; ODW 1.8.
Advertisement call. Air temperature not recorded. The call (n = 13) usually consisting of two to three consecutive notes ( Fig. 9 View FIGURE 9 A). The notes were emitted at a rate of 4.4 ± 0.4 (3.6–4.8) per second (mean ± SD [range]) within a series. The pulse group of each note usually comprised a single pulse (up to 4, median 1) and had a length of 23.0 ± 4.0 (13–32) ms ( Fig. 9 View FIGURE 9 D). Dominant frequency ranged between 3688 and 4063 Hz ( Fig. 9 View FIGURE 9 C). Although calls were not recorded under optimal breeding conditions, we consider above call parameters to be sufficiently distinct to be taxonomically informative.
Natural history. Found in grass-covered ‘dambo’ with water levels less than 30 cm in the flood plain of the Lulele River (a southern tributary draining north to the larger Congo River). Frogs were calling high up on grass and reeds. At one site, the frogs were calling at the edge of a large pool in the middle of the ‘dambo’. Other frogs recorded at the same locality included: Hyperolius adspersus , Hyperolius angolensis , Hoplobatrachus occidentalis and a unknown Kassina species.
Distribution. First reported by Laurent (1964) to occur in the area of Dundo. We recorded it further south from two more localities in the Lunda Norte Province of Angola (see Fig. 1 View FIGURE 1 ). The species is endemic to Angola, but may occur in the adjacent Democratic Republic of the Congo and possibly also northwest Zambia.
Conservation status. Currently only known from three localities (with extent of occurrence of ~3 500 km 2 and area of occupancy is ~1 500 km 2). More surveys are needed to determine the full distribution of the species, and thus allow assessment of its conservation status. At present it should be regarded as Data Deficient according to IUCN criteria (IUCN 2001). Potential threats to this species include habitat destruction due to alluvial diamond mining.
H. cinereus H. cinereus H. raymondi sp. nov. H. raymondi sp. nov. (Male n = 35) (Female n = 10) (Male n = 18) (Female n = 1)......continued on the next page Etymology. The species is named in honour of Raymond F. Laurent who first recorded the species (as H. cinereus, Laurent 1964 ) in Angola, in recognition of his many contributions to Central African herpetology.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |