
Synchaeta tremula, Muller, 1786, Muller, 1786
|
publication ID |
https://doi.org/10.11646/zootaxa.4276.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:7FC12089-2104-4893-B8A9-BE23CA19EFA7 |
|
DOI |
https://doi.org/10.5281/zenodo.6029925 |
|
persistent identifier |
https://treatment.plazi.org/id/4606AF6B-CE4C-5C41-6CAA-F94AFE58FE79 |
|
treatment provided by |
Plazi |
|
scientific name |
Synchaeta tremula |
| status |
|
S. tremula (Müller, 1786) View in CoL
Distribution and habitat. Individuals of S. tremula were found in monthly samples (each n = 3) from March to May 2016 in the plankton of both Schlossteich (ST) and Löschteich (LT). No males were observed. Both of these permanent freshwater ponds exhibit steady water in- and outflows and presented similar conditions over this time period, with a temperature of 13.2 ± 3.4 °C, a pH of 7.8 ± 0.1, an oxygen content of 94.7 ± 11.0% and 10.3 ± 0.5 mg /l, and a conductivity of 438.0 ± 83.6 µS/cm.
Specimens for LM deposited at the Natural History Museum of Berlin. Generalkatalog Freilebende Würmer , ZMB 11585 a–c.
Morphological analysis. The following morphological descriptions are of mature, parthenogenetic females of S. tremula .
Measurements. Total body length including toes: 257.0 ± 30.0 µm (range: 200–290 µm), n = 10
Foot length: 55.5 ± 6.9 µm (range: 40–65 µm), n= 10
Toe length: 12.6 ± 2.8 µm (range: 10–15 µm), n = 10
Body width at the level of the auricles: 144.0 ± 19.8 µm (range: 105–175 µm), n = 10
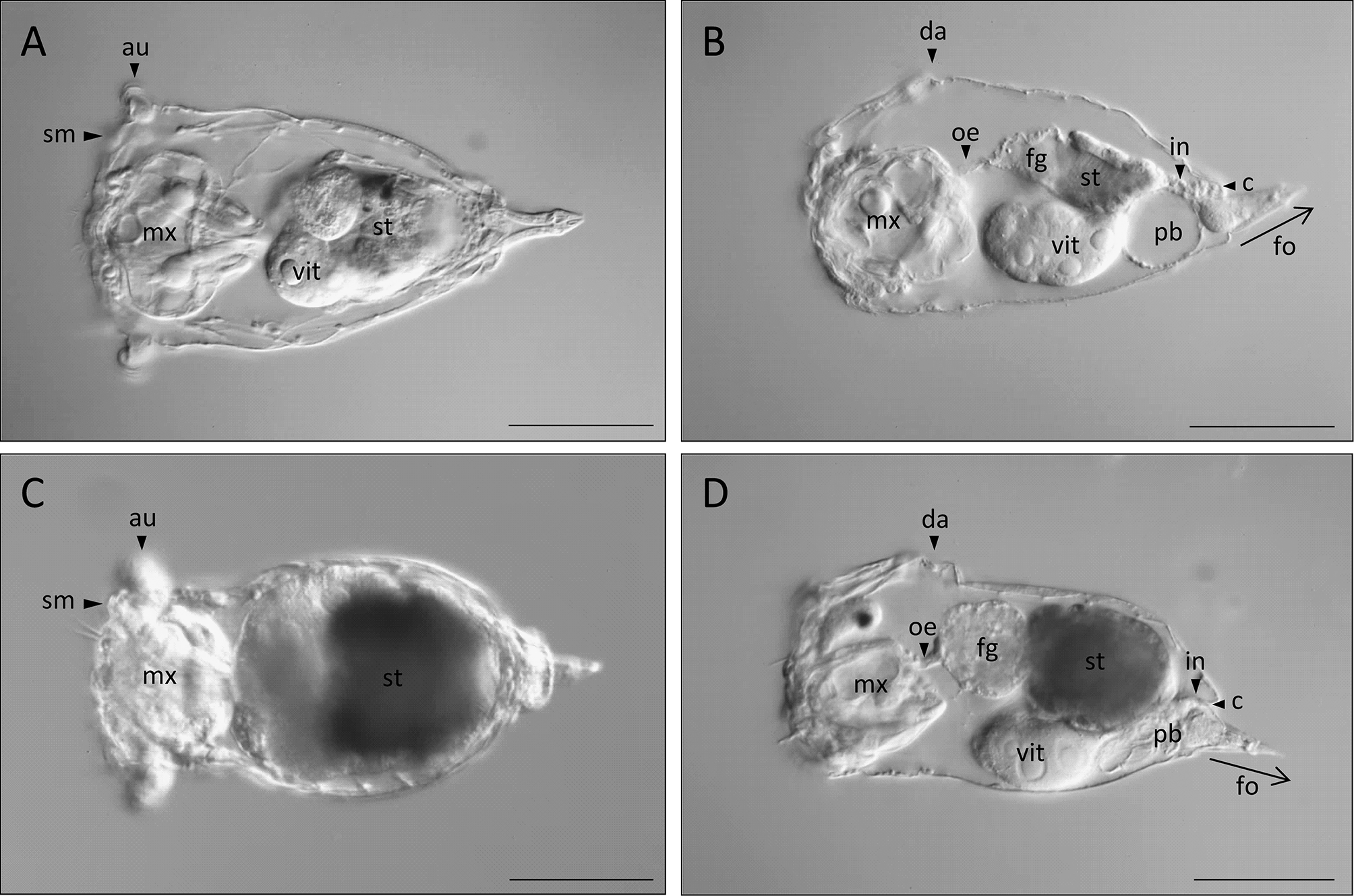
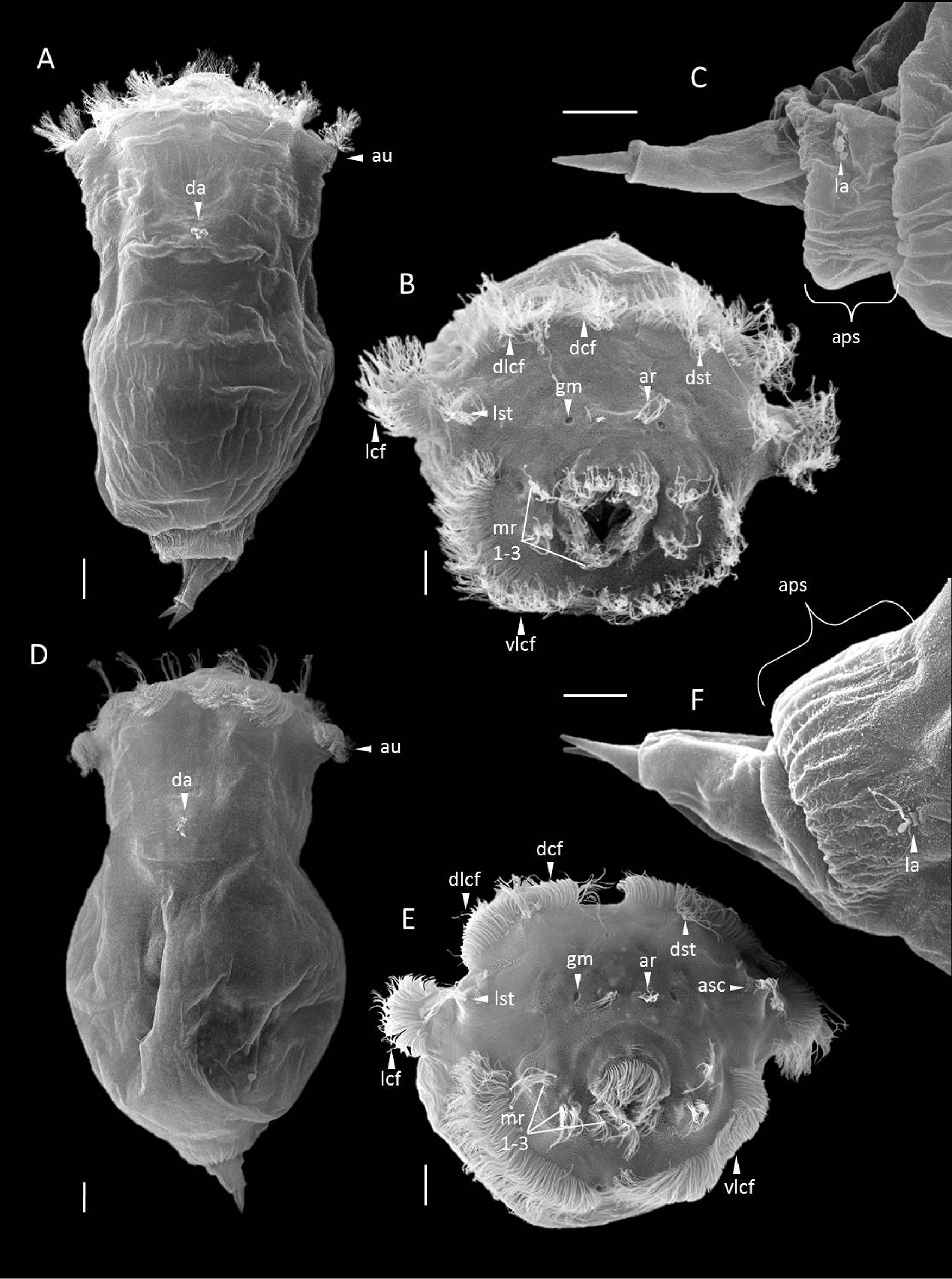
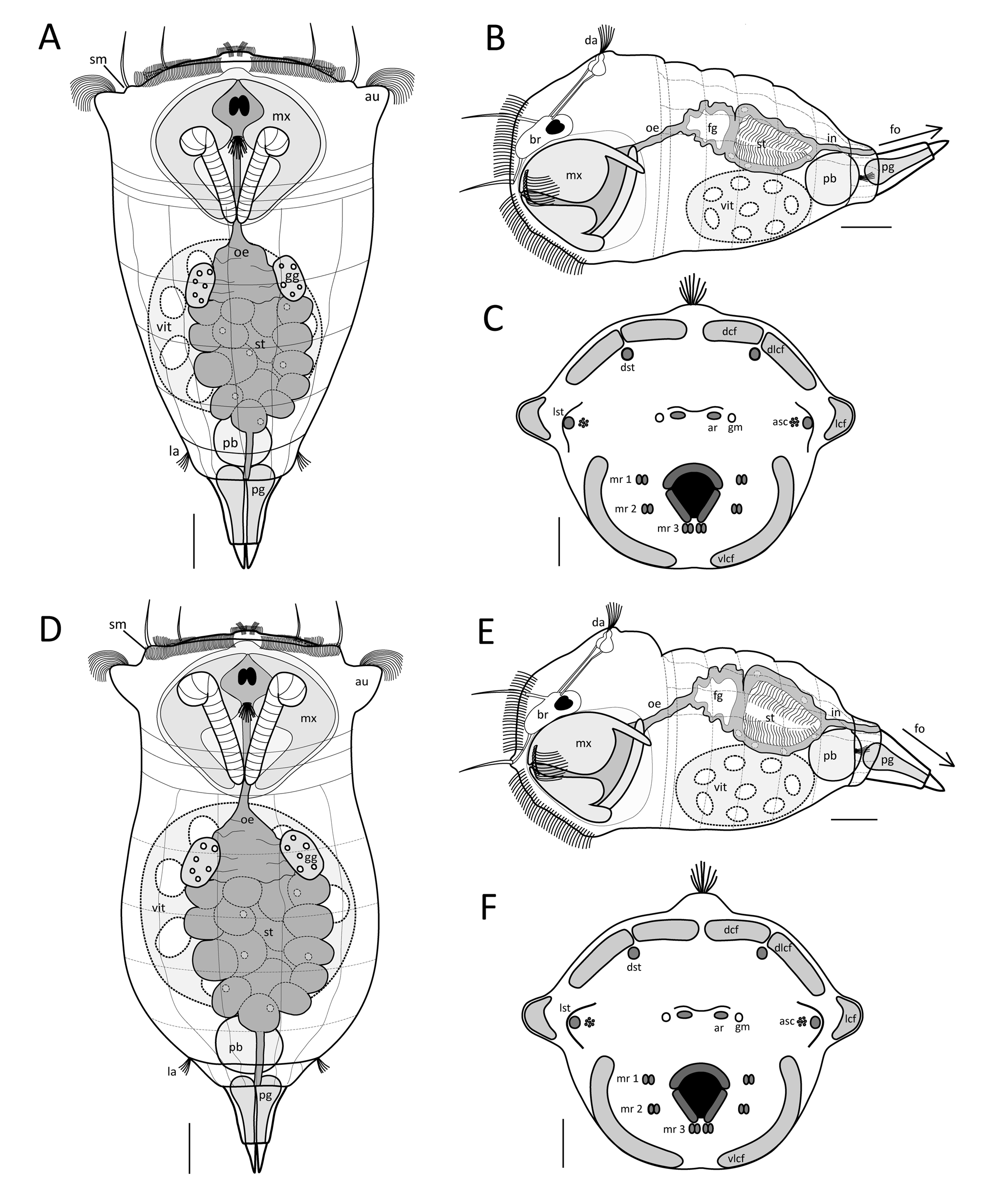
Habitus. S. tremula exhibits a flexible epidermis. The body can be subdivided broadly into the head and neck region, the trunk region, and the foot and toes. The neck is not constricted and shows a gradual transition into the trunk region, making the head region not distinctly demarcated. The overall body shape is conical, being widest in the head region at the level of the auricles and gradually narrowing caudally. The apical field is flat or very slightly convex. Each dorsolateral style on the apical field is raised on a shallow mount ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 A; sm). The auricles are small, laterally directed and not clearly protruding ( Fig. 1 View FIGURE 1 A; au). Several slight transversal folds are present in the neck region. In the head region, the apical part of the mastax shows a slight yellow to orange tone ( Fig. 1 View FIGURE 1 A; mx). The trunk region is entirely filled by the internal organs, with the stomach uniquely exhibiting a green and yellow to brownish colour ( Fig. 1 View FIGURE 1 A, B; st). The remaining body of S. tremula is almost transparent. The integument of the trunk features a few weak longitudinal and transversal striations dorsally. The distal part of the trunk presents a barely accentuated anal-pseudosegment that itself bears two lateral antennae ( Fig. 2 View FIGURE 2 C; aps, la). The conical foot is up to one-fifth of the overall body length and bends dorsally ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 B; fo). The toes are about one-quarter to one-fifth of the overall foot length including the toes, with a shape varying continuously among specimens from conical and blunt to pointed and sharp, but never curving outwardly. The entire foot is filled by separated and paired pedal glands of equal size ( Fig. 3 View FIGURE 3 A; pg). The latter are voluminous proximally and decrease in capacity distally before ending in the toes.
Rotatory organ. The rotatory organ is composed of eight ciliary fields: two dorsal, two dorsolateral, two lateral and two ventrolateral ones ( Figs. 2 View FIGURE 2 B & 3 C; dcf, dlcf, lcf, vlcf). Each of the dorsal as well as each of the ventrolateral ciliary fields are separated from one other by a narrow gap. The gap between each of the dorsal and each of the dorsolateral ciliary fields is even narrower, creating an impression of continuous dorsal cilia bands. Each lateral ciliary field is distinctly separated from each dorsolateral and ventrolateral ciliary field and is located on laterally directed ear-like prominences, the auricles ( Figs. 1 View FIGURE 1 & 2 View FIGURE 2 A; au). On the apical field, numerous sensory receptors are present, including two dorsal and two dorsolateral sensory styles ( Figs. 2 View FIGURE 2 B & 3 C; dst, lst). Each dorsal stylus is situated ventrally to the gap between each dorsal and dorsolateral ciliary field, whereas each dorsolateral stylus is located next to each auricle between the dorso- and ventrolateral ciliary fields. The latter styles are raised on shallow prominences on which a cluster of ampulla shaped cilia is present ( Fig. 3 View FIGURE 3 C; asc). In the centre of the apical field, two distinctly separated apical receptors are present that are each composed of a tuft of sensory cilia ( Figs. 2 View FIGURE 2 B & 3 C; ar). A gland meatus is located laterally to each apical receptor ( Figs. 2 View FIGURE 2 B & 3 C; gm). In the ventral half of the apical field, three pairs of ciliated receptors are found on each side of the mouth ( Figs. 2 View FIGURE 2 B & 3 C; mr 1–3). Two vertically arranged pairs are located laterally to the mouth, with a third pair situated ventrolaterally to it. A thin ciliary band adjoins to the mouth dorsally and laterally.
Nervous and sensory systems. The cerebral ganglion is situated centrally in the dorsal head region and contains two cerebral eyes ( Fig. 3 View FIGURE 3 B; br) that are slightly separated or incompletely fused and are of equal or marginally unequal size ( Fig. 3 View FIGURE 3 A). Pigment granules, when present, are accumulated at the cerebral eyespots. Frontal eyespots were never observed. On the apical field, two dorsal and two dorsolateral tactile styles are present that are each composed of several long sensory cilia. The base of each dorsal stylus is not thickened, but slightly so in each dorsolateral stylus, where it constitutes a shallow mount ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 A; sm). Each stylus is connected to the brain by a single cerebral nerve. The apical receptors ( Figs. 2 View FIGURE 2 B & 3 C; ar) are paired and distinctly separated, with each one comprising a tuft of cilia that arises from a sub-surfaced pocket and is connected to the brain by a single cerebral nerve. Each pair of ciliated mouth receptors ( Figs. 2 View FIGURE 2 B & 3 C; mr 1–3) is composed of two slightly separated tufts of sensory cilia that are raised on shallow prominences. The immovable and unpaired dorsal antenna is centrally located on a prominence in the neck region caudal to the cerebral eye ( Figs. 2 View FIGURE 2 A & 3 B; da). It is composed of a tuft of dorsally oriented sensory cilia that project through a circular pore in the integument that itself is surrounded by a low epidermal fold. The base of the sensory cilia arises from a sub-surfaced pocket from which two nerves connect the dorsal antenna with the brain. Paired immovable lateral antennae are present, each located laterally in the centre of the anal-pseudosegment ( Fig. 2 View FIGURE 2 C; la). For each antenna, a tuft of caudolaterally oriented sensory cilia projects through a pore in the integument that is likewise surrounded by a low epidermal fold. The base of each ciliary tuft also arises in a sub-surfaced pocket and is connected to a single nerve.
Digestive system. The digestive tract is subdivided into the mouth, mastax, oesophagus, foregut, stomach, intestine and the cloaca. The apical mouth is located below the longitudinal axis of the body, is oriented slightly ventrally and opens directly into the mastax cavity ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 B). It is surrounded dorsally and laterally by ciliary bands ( Fig. 2 View FIGURE 2 B). The mastax is a functional unit comprising the extensive trophus elements together with a complex muscular apparatus that interconnects the mouth with the oesophagus ( Fig. 1 View FIGURE 1 A, B; mx). It occupies the entire head and neck region in its length but not entirely in its width. The virgate trophus is specifically of the S. tremula - type, the morphology of which is described in the section “Trophi of S. tremula and S. tremuloida ”. The short oesophagus originates from the dorsal mastax region and is extended in its caudal half to form a contractile foregut that inserts itself into the dorsally situated and large-celled stomach ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 B; fg, oe, st). The inner wall of the mostly yellow to brownish coloured stomach is densely covered by long cilia and peristaltic movements can be observed frequently in live individuals ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 A, B; st). At the level of the transition from the foregut to the stomach, a pair of ovate to kidney-shaped gastric glands is present, one on each side of the digestive tract ( Fig. 3 View FIGURE 3 A; gg). Caudally, a distinct constriction is present in the transition from the stomach to the intestine. The intestine is very short and connects the stomach to the similarly very short cloaca ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 B; in) that opens dorsally to the environment at the distal end of the anal-pseudosegment.
Reproductive system. The unpaired, large vitellarium is located ventrally in the trunk region and exhibits eight large nuclei ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 A, B; vit).
Excretory system. The protonephridial bladder is located ventrally in the distal part of trunk and opens into the cloaca ( Figs. 1 View FIGURE 1 & 3 View FIGURE 3 B; pb). Contractile movements can be observed frequently in live individuals.
Swimming behaviour. Individuals are pelagic and swim in a straightforward motion with a quick rotation about the longitudinal axis. They also adhere themselves frequently to objects (e.g. plants) via a mucus thread secreted by the pedal glands, combined with a quick twisting movement about their longitudinal axis. Diagnosis. See Table 1.
S. tremula S. tremuloida Distribution and habitat. Individuals of S. tremuloida were found in monthly samples (each n = 3) from November 2015 to January 2016 in partially frozen, temporary puddles at Haarenniederung (HN) and Haarenstau (HS). Again, no males were observed. Both habitats exhibit no steady water in- or outflow, contain a high amount of decomposing organic matter, and presented similar conditions with a temperature of 6.9 ± 2.2 °C, a pH of 6.30 ± 0.07, a low oxygen content of 40.0 ± 3.0 % and 3.9 ± 0.9 mg /l, and a low conductivity of 230.0 ± 17.7 µS/cm.
Specimens for LM and SEM deposited at the Natural History Museum of Berlin. Generalkatalog Freilebende Würmer , ZMB 11586 a–e.
Morphological analysis. The following morphological descriptions are of mature, parthenogenetic females of S. tremuloida .
Measurements. Total body length including toes: 274.5 ± 22.5 µm (range: 245–325 µm), n = 10
Foot length: 52.0 ± 5.4 µm (range: 45–65 µm), n = 10
Toe length: 15.5 ± 1.6 µm (range: 15–20 µm), n = 10
Body width at the level of the auricles: 139.0 ± 13.2 µm (range: 110–155 µm), n = 10
Habitus. S. tremuloida exhibits a flexible epidermis. The body can be subdivided into the head and neck region, the trunk region, and the foot and toes. The neck region is constricted and delimits the head from the trunk region. The overall body is bell-shaped, with its widest expansion at the levels of the auricles and of the anterior trunk region. The apical field is slightly convex. Each dorsolateral style on the apical field is raised on a shallow- or medium-sized mount ( Figs. 1 View FIGURE 1 C & 3 D; sm). The auricles are of medium size, directed laterally, and clearly protruding ( Figs. 1 View FIGURE 1 C & 3 D; au). Several slight transverse folds are present in neck region. In the head region, the apical part of the mastax can show a slight yellowish tone ( Fig. 1 View FIGURE 1 C; mx). The trunk is as wide as the head region at the level of the auricles and decreases abruptly in width moving caudally. It is entirely filled by the internal organs, with the large stomach being distinctly emphasised by its green-yellow to brown colour ( Fig. 1 View FIGURE 1 C, D; st). The integument exhibits a few weak longitudinal and transverse striations dorsally. A distinct analpseudosegment is present caudally, bearing two lateral antennae located laterally within the crease between the caudal trunk region and the pseudosegment itself ( Fig. 2 View FIGURE 2 F; aps, la). The conical foot is about one-fifth of the overall body length and bends ventrally ( Figs. 1 View FIGURE 1 D & 3 E; fo). The toes are up to one-quarter of the total foot length including the toes, pointed and sharp, but never curved. The entire foot is filled by paired and separated pedal glands of equal size ( Fig. 3 View FIGURE 3 D; pg). The latter are voluminous proximally and narrow distally before the foot ends in the toes.
Rotatory organ. The rotatory organ is composed of eight ciliary fields: two dorsal, two dorsolateral, two lateral and two ventrolateral ones ( Figs. 2 View FIGURE 2 E & 3 F; dcf, dlcf, lcf, vlcf). Each of the dorsal and each of the ventrolateral ciliary fields are separated from one other by a narrow gap. A very narrow gap is present between the dorsal and dorsolateral ciliary fields, which conveys an impression of a continuous ciliary band. Each lateral ciliary field is distinctly separated from each of the dorsolateral and ventrolateral fields and is located on laterally directed and protruding prominences ( auricles; Fig. 2 View FIGURE 2 D; au). On the apical field, numerous sensory receptors are present, including two dorsal and two dorsolateral sensory styles ( Figs. 2 View FIGURE 2 E & 3 F; dst, lst). Each dorsal stylus is situated ventrally to the gap between the dorsal and dorsolateral ciliary fields, whereas each dorsolateral stylus is located next to each auricle between the dorso- and ventrolateral ciliary fields. The dorsolateral styles are raised on shallow to medium prominences on which a cluster of ampulla shaped cilia is present ( Figs. 2 View FIGURE 2 E & 3 F; asc). In the centre of the apical field and dorsal to the mouth, two distinctly separated apical receptors are present that are each composed of a tuft of sensory cilia ( Fig. 2 View FIGURE 2 E & 3 F; ar). A gland meatus is situated lateral to each apical receptor ( Figs. 2 View FIGURE 2 E & 3 F; gm). In the ventral half of the apical field, three pairs of ciliated receptors are found on each side of the mouth, with two pairs of vertically arranged receptors located laterally to the mouth and one pair ventrolaterally to it ( Figs. 2 View FIGURE 2 E & 3 F; mr 1–3). A thin ciliary band encircles the mouth dorsally and laterally.
Nervous and sensory systems. The cerebral ganglion is situated centrally in the dorsal head region and contains the cerebral eyes ( Fig. 3 View FIGURE 3 E; br). These two red eyespots are of equal or marginally unequal size and fused anteriorly ( Fig. 3 View FIGURE 3 D). Pigment granules, when present, are accumulated at the cerebral eyespots, whereas frontal eyespots were never observed. Two dorsal and two dorsolateral styles are present on the apical field, each composed of several long sensory cilia ( Fig. 2 View FIGURE 2 E; dst, lst). The base of each dorsal stylus is not thickened, whereas it is slightly thickened in each dorsolateral stylus to form a shallow to medium sized mount ( Figs. 1 View FIGURE 1 C & 3 D; sm). Each stylus is connected to the brain by a single cerebral nerve. On the apical field, the paired apical receptors are distinctly separated from one other ( Figs. 2 View FIGURE 2 E & 3 F; ar). Each receptor comprises a tuft of sensory cilia, the base of which results in a sub-surfaced pocket. Each receptor is likewise connected to the brain by a single cerebral nerve. The mouth is surrounded by two pairs of vertically arranged mouth receptors laterally as well as another pair ventrolaterally ( Figs. 2 View FIGURE 2 E & 3 F; mr 1–3). Each receptor pair is composed of two slightly separated tufts of sensory cilia that are raised on shallow prominences. One immovable and unpaired dorsal antenna is located centrally on a distinct prominence in the neck region caudal to the cerebral eye ( Figs. 1 View FIGURE 1 D & 3 E; da). It consists of a tuft of dorsally oriented sensory cilia that projects through a circular pore in the integument that itself is surrounded by a low epidermal fold. The base of the sensory cilia is found in a sub-surfaced pocket from which two nerves connect the dorsal antenna with the brain. Two immovable lateral antennae are present, each located laterally within the crease between the caudal trunk region and the anal-pseudosegment ( Fig. 2 View FIGURE 2 F; la). Each lateral antenna is composed of a tuft of caudolaterally oriented sensory cilia projecting through separate pores in the integument that, in turn, are surrounded by low epidermal folds. The base of each tuft of cilia is found in a sub-surfaced pocket and is joined to a single nerve.
Digestive system. The digestive system is subdivided into the mouth, mastax, oesophagus, foregut, stomach, intestine and the cloaca. The apical mouth is located below the longitudinal axis of the body and is surrounded dorsally and laterally by thin ciliary bands ( Fig. 2 View FIGURE 2 E). It is oriented slightly ventrally and opens directly into the mastax cavity. The very large mastax is composed of the extensive trophus elements together with a complex muscular apparatus ( Fig. 1 View FIGURE 1 C, D; mx) and takes up the entire head and neck region in both its length and width. The virgate trophus of S. tremuloida is specifically of the S. tremula - type, the morphology of which is described in the section “Trophi of S. tremula and S. tremuloida ” (see below). The short oesophagus originates from the dorsal mastax region and extends into a contractile foregut that itself inserts into the dorsally situated stomach ( Figs. 1 View FIGURE 1 D & 3 E; fg, oe, st). Through its yellow-green to brownish colour, the stomach presents a conspicuous structure within the trunk ( Figs. 1 View FIGURE 1 C, D; st). Its inner wall is densely covered by long cilia and peristaltic movements are observed regularly in live individuals. At the level of the transition from the foregut to the stomach, two ovate to kidney-shaped gastric glands are present, one on each side of the digestive tract ( Fig. 3 View FIGURE 3 D; gg). Caudally, a distinct constriction separates the stomach from the short intestine that connects the stomach with the very short cloaca ( Figs. 1 View FIGURE 1 D & 3 E; c, in) that ends dorsally at the distal end of anal-pseudosegment.
Reproductive system. The large and unpaired vitellarium is ventrally located in the trunk region and exhibits eight large nuclei ( Figs. 1 View FIGURE 1 D & 3 E; vit).
Excretory system. The protonephridial bladder is ventrally located in the distal part of trunk and leads to the cloaca ( Figs. 1 View FIGURE 1 D & 3 E; pb). Contractile movements can be observed frequently in live individuals.
Swimming behaviour. Individuals swim with a straight to slightly coiled motion, rotating thereby about the longitudinal axis. The swimming behaviour is characterised by many sudden directional changes. Individuals often adhere themselves to objects (e.g. organic matter) via a mucus thread secreted by the pedal glands, combined with a slow twisting movement around their longitudinal axis.
Diagnosis. See Table 1.
| ZMB |
Museum f�r Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |