
Halecium labrosum Alder, 1859
|
publication ID |
https://doi.org/ 10.1080/00222930400001319 |
|
persistent identifier |
https://treatment.plazi.org/id/4B6087F1-8B5B-FFDF-CB8C-FF0CFBB5FF67 |
|
treatment provided by |
Carolina |
|
scientific name |
Halecium labrosum Alder, 1859 |
| status |
|
Halecium labrosum Alder, 1859 View in CoL
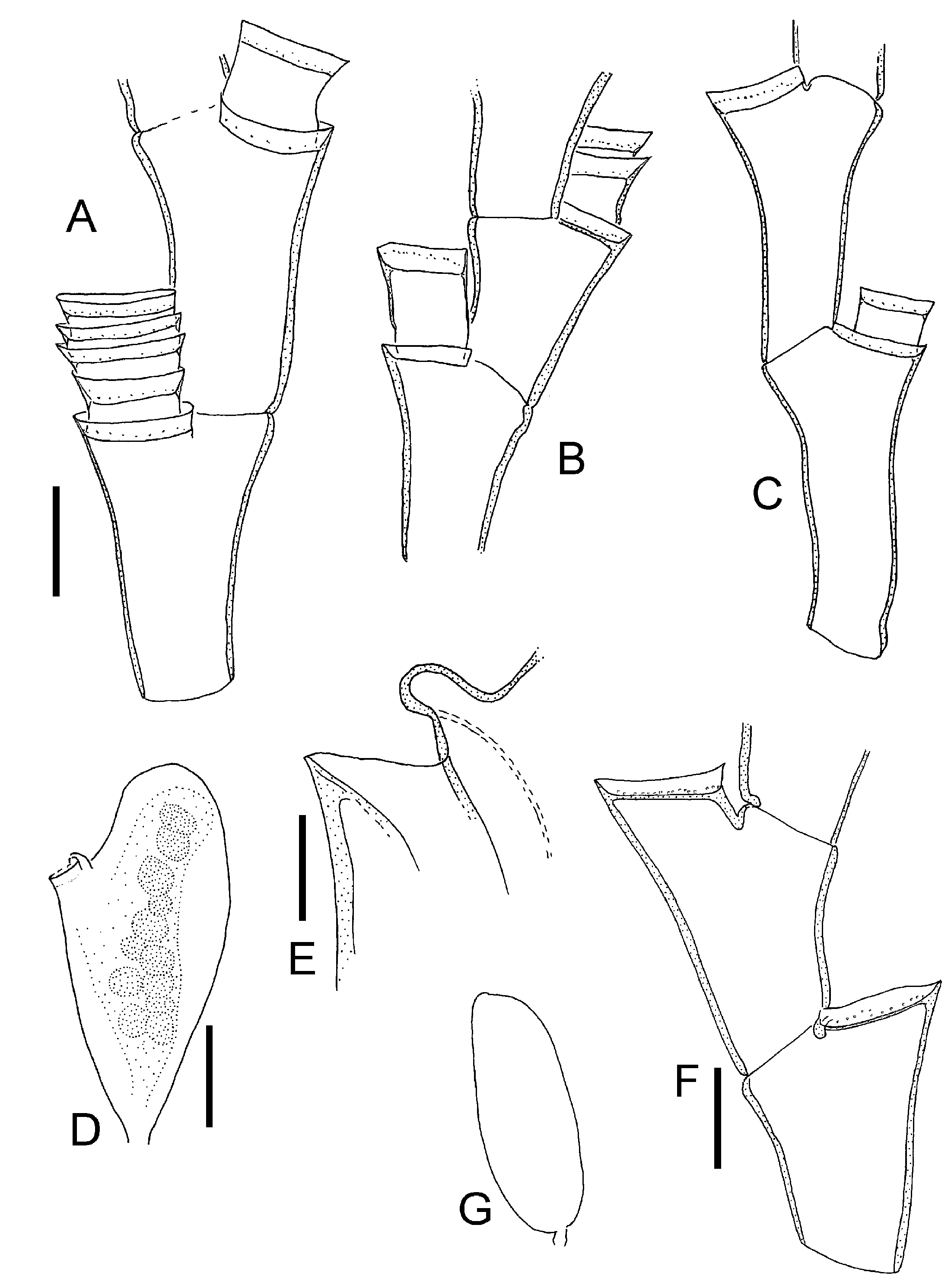
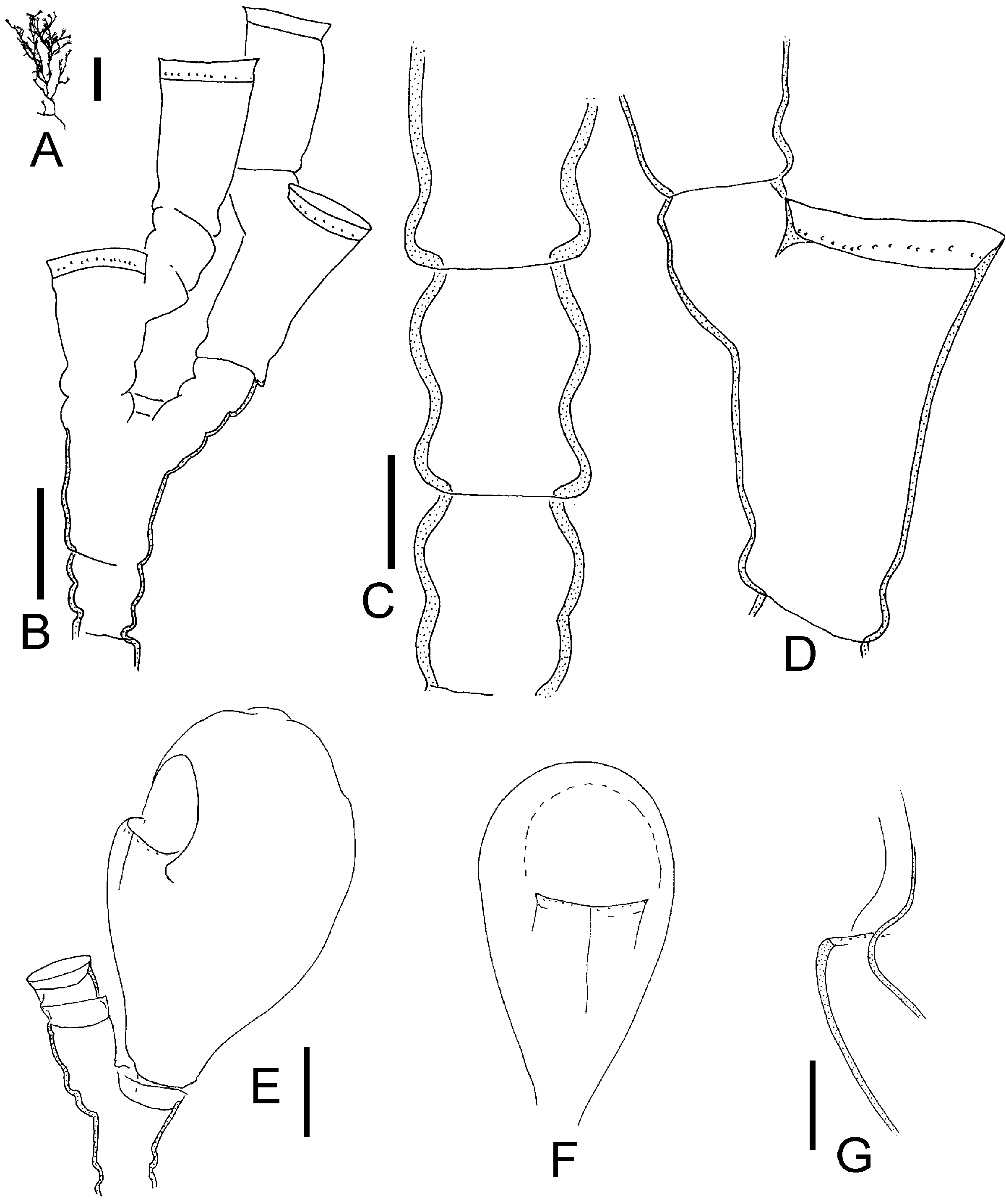
( Figures 9 View Figure 9 , 10 View Figure 10 )
Halecium labrosum Alder 1859, p 354 View in CoL , Plate 13; Hincks 1868, p 225, Figure 27, Plate 44 Figure 1 View Figure 1 ; Levinsen 1893, p 204, Plate 8 Figures 8 View Figure 8 , 9 View Figure 9 ; Broch 1910, p 148, Figures 7 View Figure 7 , 8 View Figure 8 , Plate 2 Figure 4 View Figure 4 ; Broch 1918, p 45, Figure 19; Naumov 1969, p 489, Figures 16B, 343, Plate 16 Figure 2 View Figure 2 ; Calder 1970, p 1506, Plate 1 Figures 6–8 View Figure 6 View Figure 7 View Figure 8 ; Cornelius 1975, p 396, Figure 7 View Figure 7 ; Cornelius and Garfath 1980, p 282; Cornelius 1995, p 282, Figure 64; Schuchert 2001, p 79, Figure 65A–D.
Halecium crenulatum Hincks 1874, p 150 , Plate 8 Figures 21–23; Levinsen 1893, p 204, synonym.
? Not Halecium reflexum Stechow 1919, p 37 View in CoL , Figures G, H.
Halecium schneideri: Leloup 1952, p 144 View in CoL , Figure 78A1–A3.
Material examined (see also Schuchert 2001)
MHNG INVE 28451 , Greenland, Holsteinsborg , 19 July 1953 . MHNG INVE 34235 , France, Brittany, Roscoff, between Islands of Astan et Ty Saozon , depth 5–10 m according to map, 21 April 1910,? leg. M. Bedot , juveniles up to 2 cm, no gonothecae. MHNG INVE 33581 ( BIOFAR 350 ), The Faroes , 62.26 ° N, 7.99 ° W, 107 m, 22 July 1988 GoogleMaps . MHNG INVE 33583 ( BIOFAR 553 ), The Faroes , 61.83 ° N, 6.32 ° W, 92 m, 22 September 1989 GoogleMaps . MHNG INVE 33524 ( BIOFAR 106 ), The Faroes , 62.28 ° N, 6.8 ° W, 70 m, 24 July 1987 GoogleMaps . MHNG INVE 33563 ( BIOICE collection), Iceland, 65.78 ° N, 14.22 ° W, 28–60 m, 24 July 1991 GoogleMaps . MHNG INVE 26684 ( BIOICE station 2099), Iceland, 66.62 ° N, 18.24 ° W, 112 m, 4 July 1988 GoogleMaps .
Description
Colonies up to 10 cm, arborescent, irregularly branched, predominantly in one plane, stem and some branches polysiphonic. Internodes of unequal length within and between colonies, perisarc corrugated or smooth. Nodes alternately oblique. Hydrotheca on short hydrophore at distal end of internode like a prolongation it, hydrophore not delimited by node, rim of hydrotheca reaching just to level of distal node of segment, sometimes overtopping it. Hydrotheca short, wall distinctly recurved. Renovations frequent, secondary hydrotheca on hydrophore that is several times as long as depth of hydrotheca, walls usually corrugated. Primary hydrophore often with adcauline semi-circular perisarc thickening (pseudodiaphragm), sometimes at adcauline base of hydrophore a projecting perisarc fold ( Figure 9 View Figure 9 B–D). Hydranths with 20–24 tentacles. Gonothecae without hydranths, females ovoid, compressed by about a factor of two, with short pedicel, distal end with oval opening that may be on a shallow neck-like process. Male gonothecae similar but smaller.
Typical dimensions
Internode lengths 0.35–1.05 mm, diameter of internodes 0.16–0.26 mm, diameter of diaphragm of primary hydrotheca 0.14–0.21 mm, depth of hydrotheca 40–90 mm. Female gonotheca 1.5–2.1 mm long and 0.8–1 mm broad, male gonotheca 1–1.2 mm long.
Distribution
Arctic-boreal species, in the North Atlantic reaching south at least to Brittany and North Sea, perhaps even Spain and Azores ( Cornelius 1995). Also Northern Pacific and Japan. Type localities: Northumberland coast, Shetland, Moray Firth, UK ( Cornelius and Garfath 1980).
Remarks
Halecium labrosum has very variable internode lengths, so much so that this variability itself becomes a diagnostic character. The lengths are variable within and between colonies. In his key to the British thecate hydroids, Cornelius (1995) characterizes H. labrosum as having a wrinkled or corrugated perisarc. While some colonies indeed have such a corrugated perisarc, especially the secondary hydrophores, there were also regularly colonies with smooth perisarc among the examined material from the North Atlantic (compare Figures 9B, C View Figure 9 and 10B View Figure 10 ).
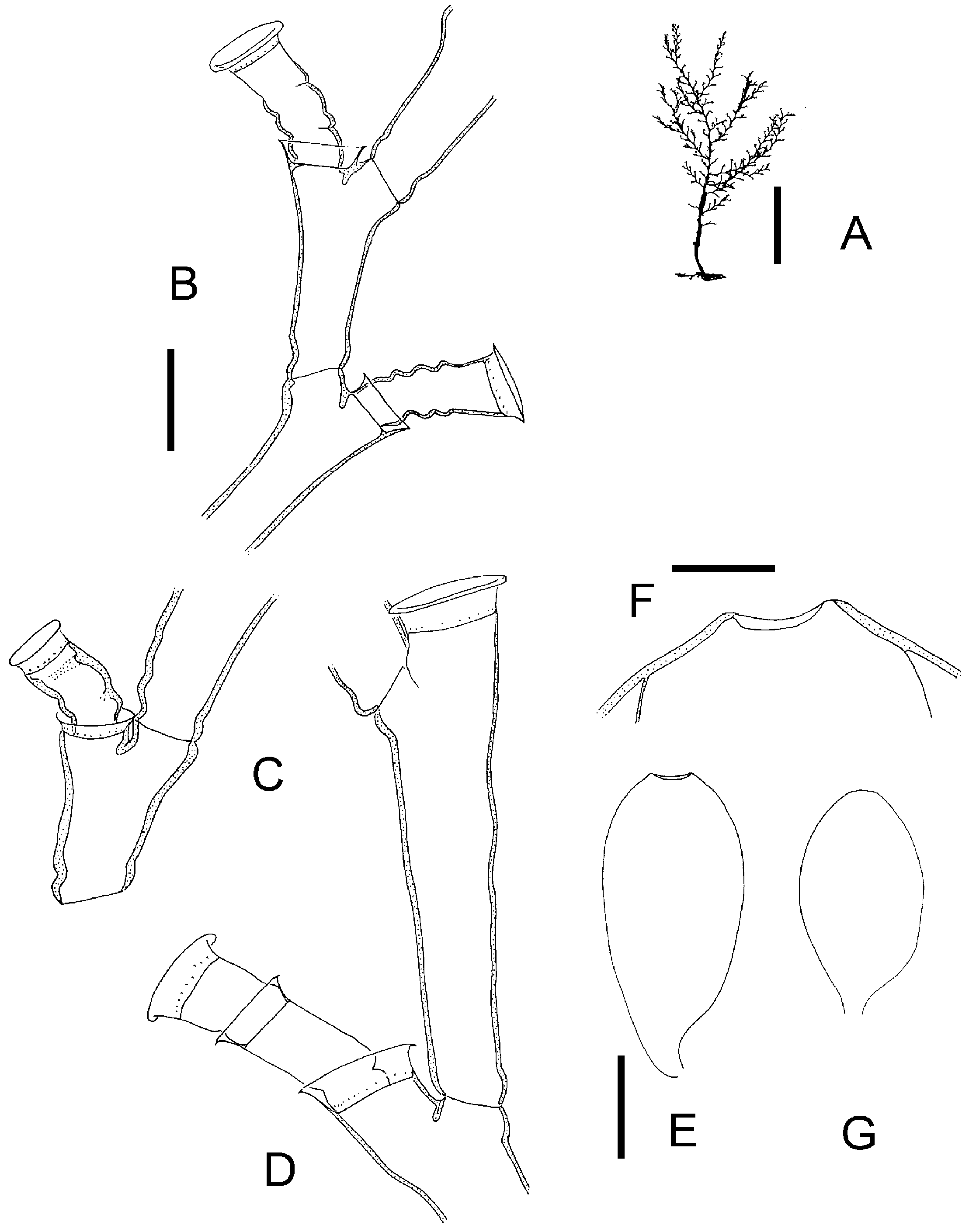
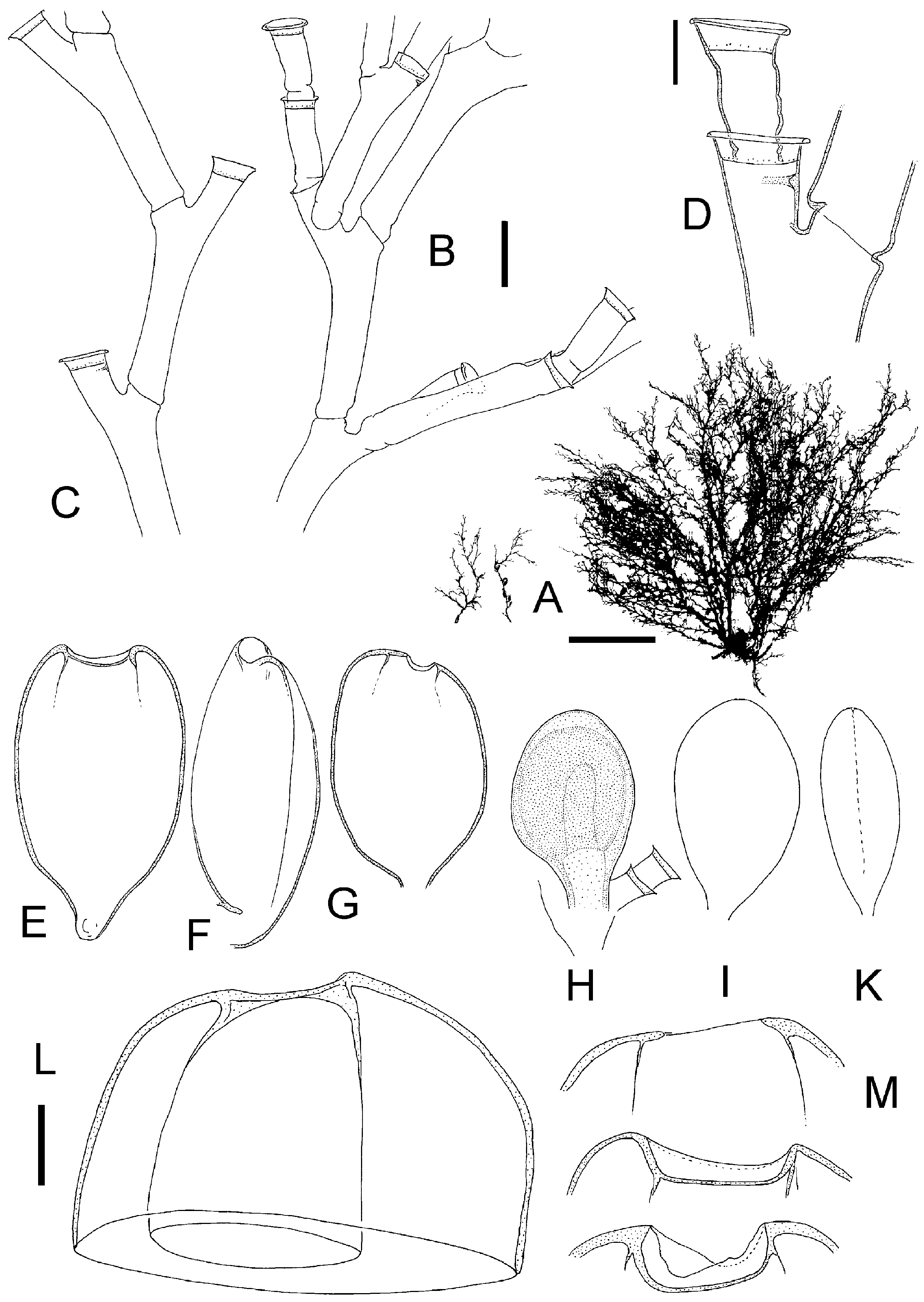
Broch (1918) considered Halecium labrosum to be an Arctic or northern Atlantic species. Likewise, Cornelius (1975) considered the English Channel as its probable southern limit. As already mentioned by Broch (1918), it is likely that at least some Mediterranean records of H. labrosum are due to a confusion with other Halecium species, notably H. mediterraneum . Halecium mediterraneum (see below) is by no means easily separable from H. labrosum and in fact might represent a southern form of the latter. The differences between the two forms are only gradual. It differs from H. labrosum in forming smaller, mostly monosiphonic shoots (but some are weakly polysiphonic!), the long hydrophore which makes the hydrotheca always overtop the distal node of the segment, the smaller diameter of the hydrotheca, and the smaller diameter of the segments (internodes). The growth forms are also different, with H. mediterraneum tending to form bushy, tangled masses, while H. labrosum is always arborescent (compare Figures 10A, E View Figure 10 and 11A View Figure 11 ).





| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halecium labrosum Alder, 1859
| Schuchert, Peter 2005 |
Halecium schneideri:
| Leloup 1952: 144 |
Halecium reflexum
| Stechow 1919: 37 |
Halecium crenulatum
| Hincks 1874: 150 |
Halecium labrosum
| Alder 1859: 354 |