
Reidnitocrella, Karanovic & Hancock, 2009
|
publication ID |
https://doi.org/10.11646/zootaxa.2324.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:55E33A9D-AB38-4FA9-9CBD-0AA24A130CE4 |
|
persistent identifier |
https://treatment.plazi.org/id/6F034DD3-613D-4661-B10C-E9AC61631A30 |
|
taxon LSID |
lsid:zoobank.org:act:6F034DD3-613D-4661-B10C-E9AC61631A30 |
|
treatment provided by |
Felipe |
|
scientific name |
Reidnitocrella |
| status |
gen. nov. |
Genus Reidnitocrella gen. nov.
Diagnosis. Medium sized Ameiridae , with cylindrical habitus and no distinct demarcation between prosome and urosome. Integument weakly chitinized and without cuticular windows; hyaline fringe of all somites smooth. First pedigerous somite incorporated into cephalothorax. Prosome weakly ornamented with moderately large sensilla, urosome with or without ventral posterior rows of small spinules. Genital doublesomite with visible suture laterally and dorsally; genital field with single large copulatory pore, wide copulatory duct and two small semicircular seminal receptacles; single small genital aperture covered by fused reduced sixth legs. Anal operculum narrow and convex, not reaching to posterior end of anal somite, unornamented. Caudal rami ovoid or cylindrical, 1.5 times as long as their greatest width and slightly divergent; dorsal seta inserted near posterior margin and very close to inner margin, about twice as long as ramus; proximal lateral seta arising somewhat dorsolaterally at middle; distal lateral seta arising at 5/6 and laterally; inner apical seta small; principal apical setae bipinnate, with breaking plane. Antennula long and slender, eight-segmented in female and ten-segmented and not strongly geniculate in male; with seta on first segment. Antenna composed of coxa, basis, two-segmented endopod and one-segmented exopod; exopod armed with three setae. Mandibula with two-segmented palp; basis unarmed, endopod with four apical setae. All swimming legs with three-segmented exopod. Endopod of first leg three-segmented, while that of second leg two-segmented, third leg one- or two-segmented and fourth leg always one-segmented. All exopodal segments of about same length; first exopodal segment of all legs without inner seta, second with; third exopodal segment of all legs with two outer spines, that of first, second and third legs without inner setae, that of fourth leg with two inner setae. First endopodal segment of first leg large, armed with one inner seta; subapical endopodal segment of second and third legs (when present) armed with one inner seta or unarmed; apical endopodal segment of second leg armed with one inner seta and one apical plumose seta, that of third leg with inner apical plumose seta and outer apical plumose spine, while endopod of fourth leg armed with single apical spine. Basis of first leg in male with transformed inner spine. Fifth leg same in both sexes, biramous but with baseoendopods fused both medially together and to somite; endopodal lobe not recognizable and without armature or ornamentation; basal outer seta arising from very long setophore; exopod a distinct segment, quadriform, small, armed with two or three slender smooth setae.
Type species. Reidnitocrella tianschanica ( Borutzky, 1972) comb. nov. [= Nitocrella tianschanica Borutzky, 1972: p.111 , figs 7/1–7/10].
Other species. Reidnitocrella borutzkyi sp. nov. [= Nitocrella tianschanica Borutzky, 1972: p. 111 , figs 7/ 11–7/16]; Reidnitocrella pseudotiaschanica ( Sterba, 1973) comb. nov. [= Nitocrella pseudotianschanica Sterba, 1973 ]; Reidnitocrella djirgalanica ( Borutzky, 1978) comb. nov. [= Nitocrella djirgalanica Borutzky, 1978 ].
Etymology. The new genus is named in honour to Dr Janet W. Reid (University of Virginia, USA), as recognition of her enormous contribution in the field of freshwater copepod taxonomy. She (with two other colleagues) also made the genus name Stygonitocrella Reid, Hunt & Stanley, 2003 available, by designating the type species. Her surname is prefixed to the existing genus name Nitocrella . Gender feminine.
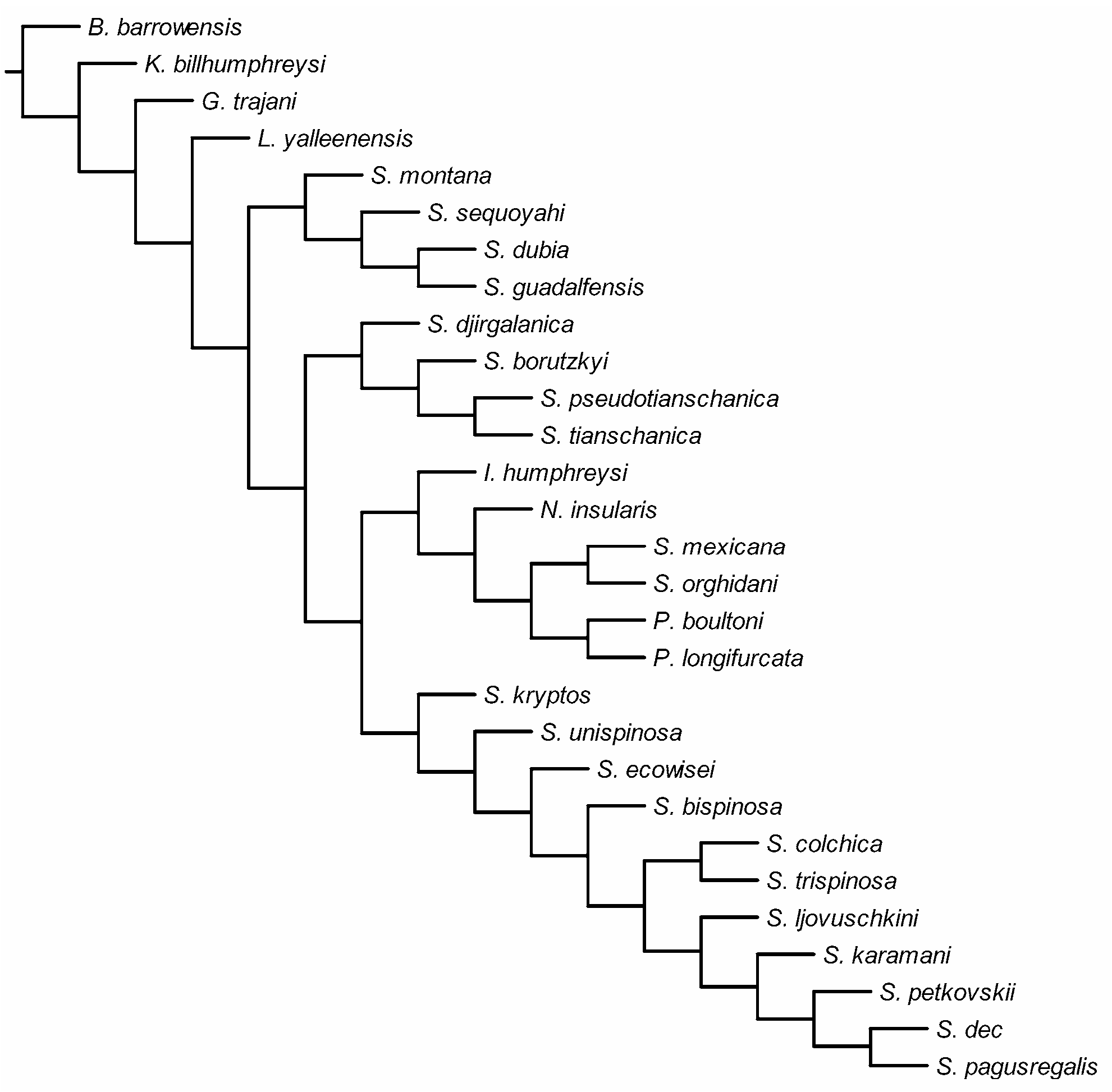
Remarks. Species of the genus Reidnitocrella gen. nov. form a well supported clade on the cladogram ( Fig. 18 View FIGURE 18 ) of this group of freshwater ameirids ( Stygonitocrella s. l.) and, interestingly, all come from a relatively small area in central Asia. Reidnitocrella pseudotianschanica ( Sterba, 1973) was described from central Afghanistan, while R. tianschanica ( Borutzky, 1972) , R. djirgalanica ( Borutzky, 1978) and R. borutzkyi sp. nov. all come from the interstitial of Lake Issyk-Kul in Kyrgyzstan (see Borutzky 1972, 1978; Sterba 1973).
Their armature formula of the ultimate endopodal segment of the second to fourth legs is either 2.2.1 or 2.1.1, as is the endopodal segmentation formula of the same legs. But it is the nature of these elements that is of far more importance. While two armature elements on the ultimate endopodal segment of the second leg is also found in Eduardonitocrella mexicana ( Suárez-Morales & Iliffe, 2005) comb. nov., Neonitocrella insularis ( Miura, 1962) and Megastygnitocrella karamani ( Petkovski, 1959) comb. nov., they represent the ancestral two apical setae in E. mexicana ( Table 2, characters 26 & 27), the outer subapical spine and outer apical seta in N. insularis and M. karamani (characters 25 & 26) and the ancestral outer apical and distal inner setae in the genus Reidnitocrella (characters 26 & 28). In fact, the nature of these elements in Reidnitocrella is the same in only two other species: Gordanitocrella trajani sp. nov. and Lucionitocrella yalleenensis sp. nov. As we discussed above, these two species have a number of unique plesiomorphic characters (and some autapomorphic) and are not closely related to any of the recent Stygonitocrella s. l. species. If, for example, we just compare the armature elements on the endopod of the third leg, it becomes clear that those of Gordanitocrella gen. nov. represent characters 36, 38 & 39, while those of Lucionitocrella gen. nov. represent characters 36, 37 & 39 and of Reidnitocrella characters 35 & 36.
On the other hand, the nature of the armature elements on the ultimate endopodal segment of the third and fourth legs is exactly the same in Reidnitocrella gen. nov. and Megastygonitocrella gen. nov., which suggests that these two genera are probably most closely related. However, all Megastygonitocrella species have the ancestral outer subapical spine on the endopod of the second leg (character 25) present (absent in Reidnitocrella ) and in all species, except in M. karamani ( Petkovski, 1959) , this is also the only armature element. Also all Megastygonitocrella species have lost the inner seta on the second exopodal segment of the first leg (character 17), while it is present in all four Reidnitocrella species. Unfortunately, the mouth appendages are practically unknown for all four representatives of the latter genus, so many potentially valuable characters could not be included in the cladistic analysis.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.