
Physiphora Fallén 1810
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4087.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C510CF71-0039-478A-91ED-BFD65B6FE0BE |
|
DOI |
https://doi.org/10.5281/zenodo.6066577 |
|
persistent identifier |
https://treatment.plazi.org/id/5101BA35-FFA6-FFF1-FF1A-E36DD46CFAA6 |
|
treatment provided by |
Plazi |
|
scientific name |
Physiphora Fallén 1810 |
| status |
|
Genus Physiphora Fallén 1810 View in CoL View at ENA
Physiphora Fallén, 1810: 11 . Type species: Chrysomyza splendida Fallén, 1817 (= Musca alceae Preyssler, 1791 ), by subsequent monotypy in Fallén, 1817: 3.
Chrysomyza Fallén, 1817: 3 . Type species: Chrysomyza splendida Fallén, 1817 (= Musca alceae Preyssler, 1791 ), by monotypy.
Cliochloria Enderlein, 1927: 103 . Type species: Musca aenea Fabricius, 1794 (= Ulidia clausa Macquart, 1843 ), by original designation.
Diagnosis. The genus possesses all the diagnostic characters of the tribe Ulidiini: veins R1 and R4+5 bare, cell cup with triangular lobe along vein A1, female abdominal tergites 4–6 without anteromedial apodemes, female tergite and sternite 6 very short and transverse, gonites bearing no setulae, strongly asymmetrical: right gonite forming projection, left gonite flat and vestigial anteroventral cornua of hypandrium connected to lateral projections of phallus (epiphallus?), phallus very large, mostly bare, with membranous extension at middle of its length (caecum) and apical part with glans, formed by modified spines or scales; epandrium and surstyli separated by seam; subepandrial sclerite (= bacilliform sclerite, decacternum) v-shaped, consisting of two anteriorly joined stripe-like sclerites more or less uniformly covered by setae, male cerci flat and not protuding posteriorly, female ovipositor with long taeniae and fine scales on eversible membrane; aculeus with moderately long, setulose and sclerotized segment 8 and oval, long setulose cercal unit; vagina with simple, bar-like ventral receptacle; 3 spherical spermathecae, one at right side and two on Y-shape bifurcated common duct at left side.
Species of Physiphora can be easily distinguished from the species of Ulidia Meigen and Timia Wiedemann by the combination of ventro-laterally opened antennal grooves, entirely greenish or bluish metallic body without long setulae, hyaline wings with long lobe of cell cup.
From superficially similar, metallic greenish species of the genus Euxesta Meigen (tribe Lipsanini), species of Physiphora can be distinguished from the combination of hyaline wings with long lobe of cell cup (usually with dark pattern and short lobe of cup in Euxesta ), scutum largely bare, with setulae restricted mostly to dorsocentral and supra-alar rows (evenle distributed over scutum in Euxesta ), phallus with glans (simple tubular phallus without glans in Euxesta ) and having 3 spermathecae (2 spermathecae in Euxesta ).
More extended comparative diagnosis is as follows: antennal grooves entirely opened ventro-laterally (in all Ulidia and Timia antennal grooves deeply impressed and sharply delimited also ventro-laterally); thorax and abdomen with strong metallic golden, green, blue or purple sheen (at most abdomen with greenish metallic sheen in some Timia ), dorsally at most sparsely or non-setulose without large microtrichose fields (usually densely and long setulose in Ulidia and Timia and often largely white microtrichose in Timia ), wing with long triangular lobe of cell cup at least as long as vein Cu2+A1 (usually with short lobe in Ulidia and most Timia ), wing entirely hyaline (except in P. meyi sp. n., possessing small dark spot at R4+5 and M junction with costa; in Ulidia and Timia often widely darkened, but dark pattern, if present, not exceeding cell r2+3); frons with pair of white microtrichose parafrontal spots at sides (frons either entirely shining or widely microtrichose in Ulidia and Timia ); compound eye green with four purple bands in live specimens (uniformly greenish or reddish in all Ulidia and Timia ); fore metatarsus often white (dark in most Ulidia and Timia , if yellow, not densely white setulose); male postabdomen with very narrow, stripe-like cerci bearing a pair of ventrally directed nipple-like projections (large and lobate in Ulidia and Timia , without such projections); spermathecae spherical, with short invaginated “necks” (in examined species of Ulidia and Timia ovoid with long “neck” deeply invaginated into spermatheca).
Description. Body mostly black, usually with strong blue, green, golden to purple or red metallic sheen ( Figs. 28 View FIGURES 28 – 30 , 53 View FIGURES 53 – 55 , 60 View FIGURES 60 – 65 , 72 View FIGURES 72 – 79 , 89 View FIGURES 89 – 96 , 138 View FIGURES 138 – 145 , 181 View FIGURES 181 – 186 , 217 View FIGURES 217 – 222 , 300 View FIGURES 300 – 305 , 342 View FIGURES 342 – 345 , 350 View FIGURES 350 – 353 , 395 View FIGURES 395 – 400 ).
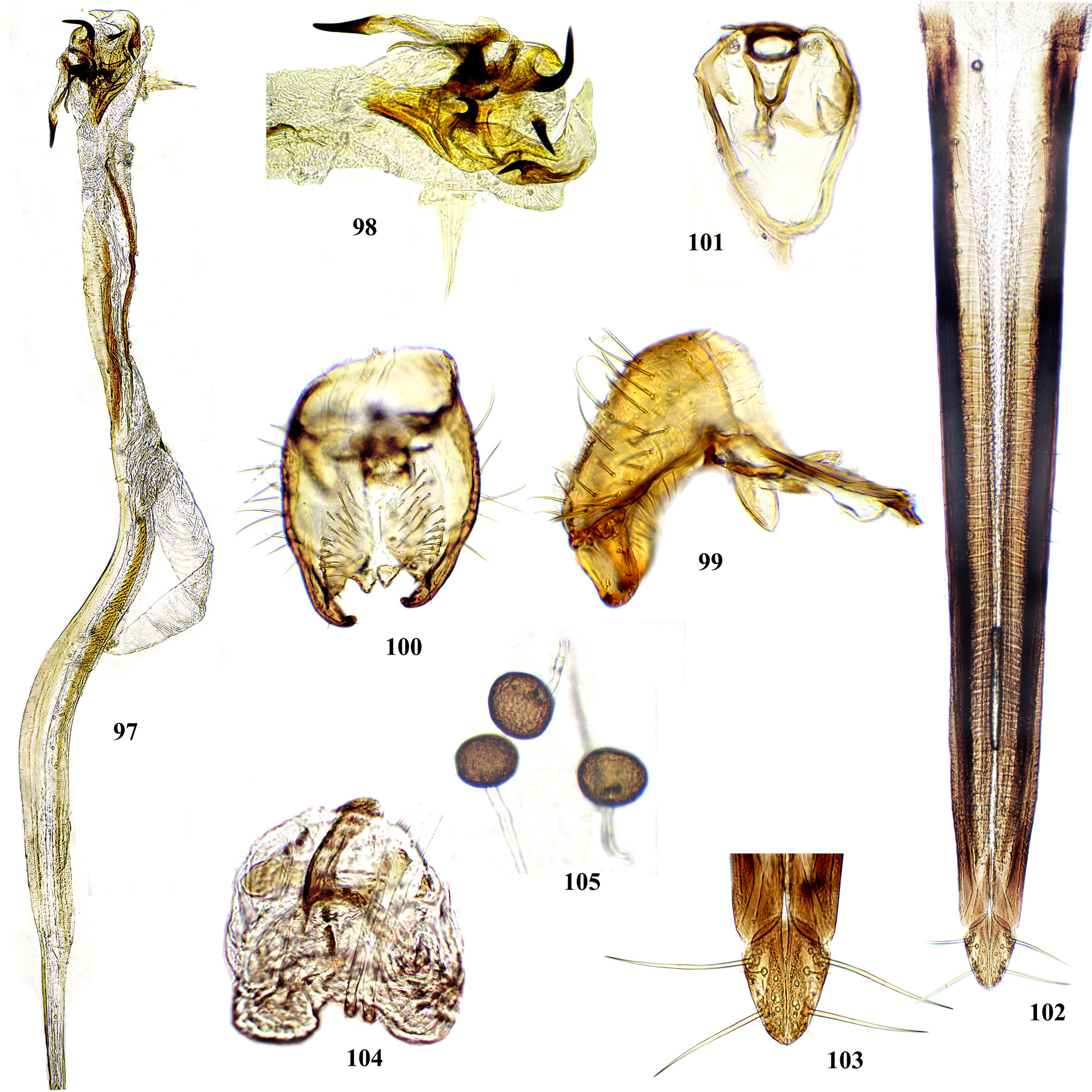
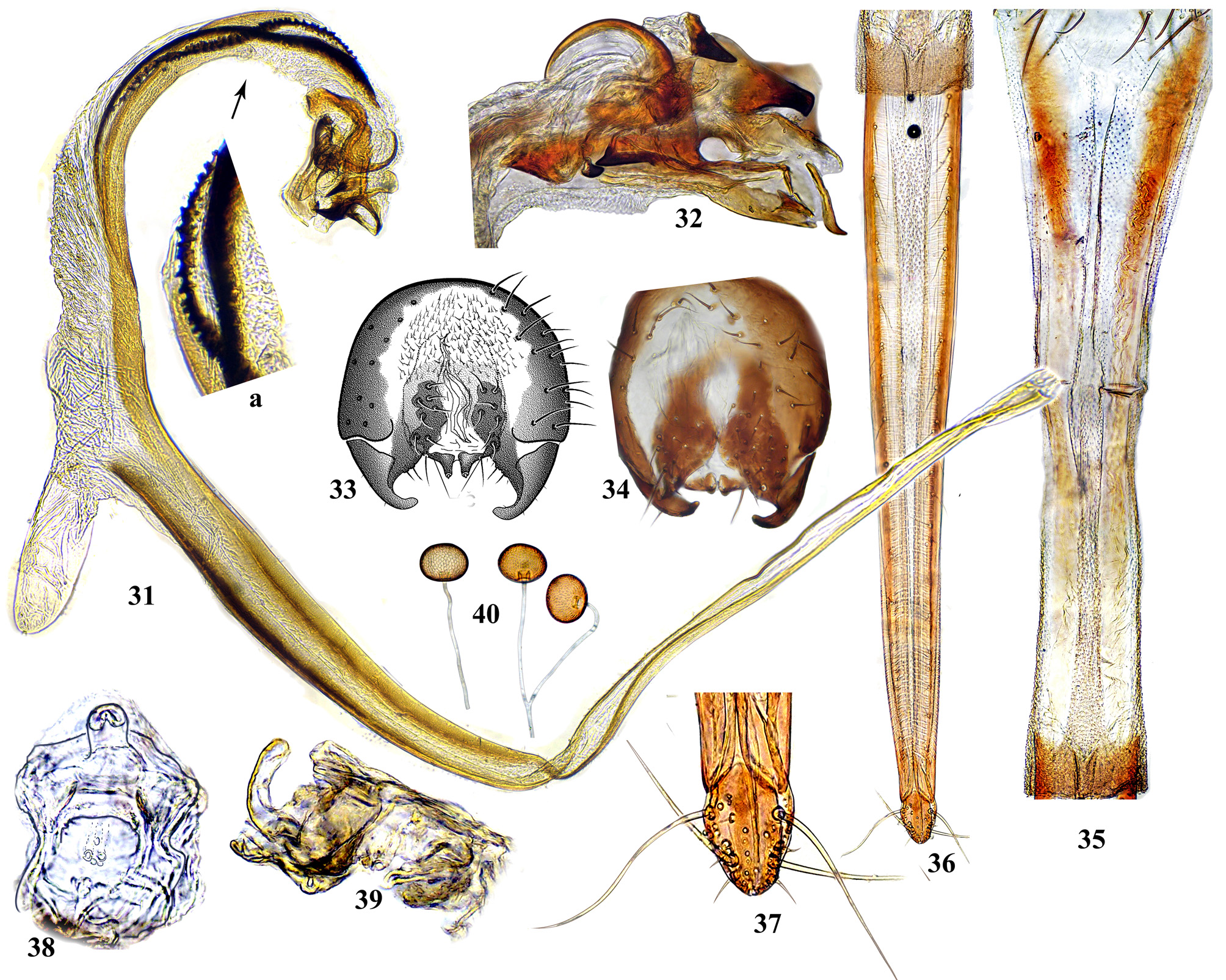
Head at most a little higher than long. Frons smooth (Figs. 13–14, 16) or rarely pitted (Figs. 15, 21, 62) or wrinkled (Figs. 22–24), entirely yellow (Fig. 19), reddish or brownish yellow with black, often greenish shining vertical plates and ocellar triangle (Figs. 11, 12, 18, 21, 58, 62) to entirely black (Figs. 10, 14, 22, 27, 44); frontal vitta usually with a few short, often inconspicuous setulae (Figs. 15, 62), in posterior part often with four low longitudinal swellings ( Figs. 29 View FIGURES 28 – 30 , 44 View FIGURES 41 – 45 , 57 View FIGURES 56 – 59 , 196 View FIGURES 194 – 199 , 220 View FIGURES 217 – 222 ), usually with a pair of spots of whitish microtrichia (Figs. 13– 18), rarely with large microtrichose areas (Figs. 10–12). Vertical plates with 2 short orbital setae; ocellar triangle with pair of short ocellar setae. Face usually saddle-like, slightly concave in profile ( Figs. 61 View FIGURES 60 – 65 , 74 View FIGURES 72 – 79 ), sometimes with wide facial carina, more or less produced anteriorly, abruptly separated from antennal grooves ( Figs. 183 View FIGURES 181 – 186 , 196 View FIGURES 194 – 199 , 352 View FIGURES 350 – 353 ); dorsal part of face either with entire white microtrichose band covering antennal grooves and facial carina (Figs. 25, 29, 73, 95, 128, 144, 170, 294, 303, 383, 397), with shallow, gray microtrichose antennal grooves, often with narrow, shining bare areas, separating λ-like white microtrichose pattern on carina from microtrichiae on grooves (Figs. 26, 43, 55, 109, 111, 208, 265, 278); ventral half of face either yellow, brown, medially or entirely black, sometimes with metallic blue or green sheen ( Figs. 55 View FIGURES 53 – 55 , 58 View FIGURES 56 – 59 , 62 View FIGURES 60 – 65 , 95 View FIGURES 89 – 96 , 220 View FIGURES 217 – 222 ). Parafacial bare or with longitudinal stripes of white microtrichia; gena bare, rarely with patches of microtrichia. Occiput mostly black, sometimes yellow or orange dorso-medially ( Figs. 89 View FIGURES 89 – 96 , 150 View FIGURES 146 – 152 , 293 View FIGURES 292 – 296 ). Pedicel short. Scape with incision, but without notch; one long pedicellar seta and 10–12 short setulae. First flagellomere oval, not over 2.2–2.7 times as long as wide, rounded at apex; arista 3-segmented. Palp flattened oval, not widened apically, usually as long and as wide as flagellomere 1, white microtrichose, with 12–15 lateral and ventral setae almost as long as palp width.
Compound eye of Physiphora golden greenish with several purplish stripes, often with two pairs of dilations medially ( Figs. 28–29 View FIGURES 28 – 30 , 56 View FIGURES 56 – 59 , 89, 94 View FIGURES 89 – 96 , 107 View FIGURES 106 – 113 , 148 View FIGURES 146 – 152 , 182–183 View FIGURES 181 – 186 , 231 View FIGURES 231 – 236 , 244, 247 View FIGURES 244 – 252 , 276, 278 View FIGURES 276 – 281 , 313, 315 View FIGURES 313 – 316 , 382–383 View FIGURES 380 – 386 ), well visible in live, freshly killed or moistured flies, but usually indistinguishable in dry specimens. Some eye patterns have been described and figured by Hendel (1909: Abb. 1–8).
Thorax entirely black, mostly with metallic greenish, copper, golden, blue or purple sheen. Postpronotal lobe with 1 seta and 3–5 setulae; proepisternum with one long and 7–8 shorter setae. Prosternum transverse, trapezoidal, without conspicuous setae. Mesonotum with scutum and postero-dorsal part of anepisternum more or less densely shagreened, from almost glossy ( Figs. 219 View FIGURES 217 – 222 , 302 View FIGURES 300 – 305 , 382 View FIGURES 380 – 386 ), sparsely rugulose and shining ( Figs. 351 View FIGURES 350 – 353 , 396 View FIGURES 395 – 400 ) to densely rugulose, subshining with metallic sheen ( Figs. 277 View FIGURES 276 – 281 , 293 View FIGURES 292 – 296 , 314 View FIGURES 313 – 316 , 324 View FIGURES 323 – 327 , 344 View FIGURES 342 – 345 ) to matt black with dull green or blue tinge ( Figs. 169 View FIGURES 167 – 173 , 264 View FIGURES 262 – 267 , 374 View FIGURES 374 – 379 ). Anterior part of mesonotal scutum and postero-dorsal part of anepisternum setose; rest of scutum usually bare, sometimes with dorsocentral and acrostichal row of very fine and short setulae ( Figs. 30 View FIGURES 28 – 30 , 63 View FIGURES 60 – 65 , 75 View FIGURES 72 – 79 , 89 View FIGURES 89 – 96 , 126 View FIGURES 124 – 130 , 142 View FIGURES 138 – 145 ) or, rarely, evenly and sparsely setulose ( Fig. 209 View FIGURES 206 – 209 ). 2 notopleural, 2 postsutural supra-alar, 1 intraalar and 1 postalar seta present. Dorsocentral and acrostichal setae very fine, inconspicuous or absent. One strong seta and 5–6 shorter setulae on posterior margin of anepisternum; katepisternum with one strong posterodorsal seta. Scutellum triangular, flattened, shagreened, with 2 pairs of scutellar setae and fine setulae on disc and margins; posteroventral margin bare or microtrichose. Subscutellum matt, sometimes short white microtrichose. Mediotergite slightly shagreened, subshining green.
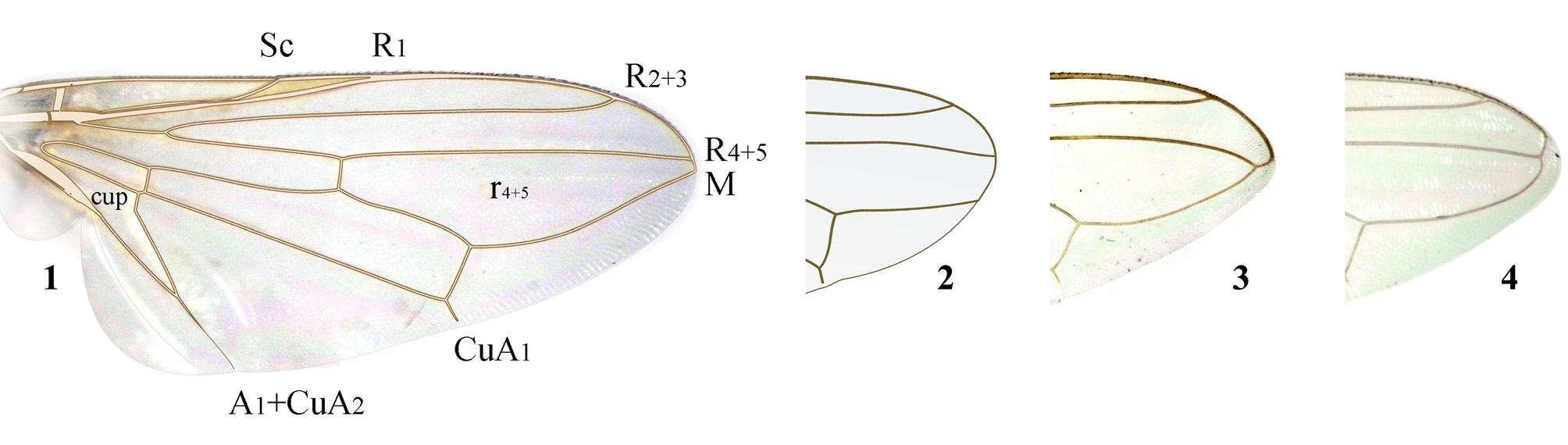
Wing entirely hyaline with yellow veins ( Figs. 1–4 View FIGURES 1 – 4 ) (rarely partly brown), in P. m ey i sp. n. with small blackish spot at apex of cell r4+5 ( Figs. 244, 245, 250 View FIGURES 244 – 252 ); vein R1 bare, cell r4+5 apically narrowed ( Figs. 1, 2 View FIGURES 1 – 4 , 185 View FIGURES 181 – 186 , 280 View FIGURES 276 – 281 ) or closed ( Figs. 64 View FIGURES 60 – 65 , 77 View FIGURES 72 – 79 , 113 View FIGURES 106 – 113 ), sometimes vein M fused with R4+5 before costal vein and wing apex, forming short petiole ( Figs. 3 View FIGURES 1 – 4 , 93, 96 View FIGURES 89 – 96 , 385 View FIGURES 380 – 386 ) or simply turning into costa before apex ( Figs. 157 View FIGURES 153 – 157 , 316 View FIGURES 313 – 316 ); cell bcu with long extension at posteroapical corner, usually as long as the remaining vein A1+CuA2 ( Figs. 1 View FIGURES 1 – 4 , 45 View FIGURES 41 – 45 , 64 View FIGURES 60 – 65 , 77 View FIGURES 72 – 79 ), not showing conspicuous variability.
Legs mostly brown or black, usually with mid and hind tarsi yellow; in P. anaglypha , P. clausa Macquart and P. flavipes Karsch femora mostly yellow and tibiae at most partly brown ( Figs. 53 View FIGURES 53 – 55 , 89 View FIGURES 89 – 96 , 146 View FIGURES 146 – 152 , 156 View FIGURES 153 – 157 ); in P. igniceps sp. n. brow in basal tree-fourths, reddish yellow apically. Fore femur posteroventrally with row of 3–9 setae, in most species conspicuously thickened and spinulose (Fig. 8). Mid leg on anterior and posterior surface with two rows of moderately long, antero- and postero-ventrally directed setae. Fore tibia, apically widened and flattened, and with brush of short and dense yellowish setae anteroapically, usually entirely brown or black colored, partly brown in P. clausa and P. flavipes (Fig. 8) or entirely yellow in P. anaglypha Séguy. Fore tarsus either entirely brown to black (Figs. 5, 76, 130, 300, 323, 342, 395) or with first segment (basitarsus, metatarsus) yellow, white setulose (Figs. 6–8, 60, 167, 182, 245, 279, 313, 350, 362, 374); in P. anaglypha fore tarsus entirely yellow (Figs. 9, 53).
Abdomen shining black, only some specimens of P. flavipes with yellow basal and apical tergites ( Fig. 153 View FIGURES 153 – 157 ); tergites often finely shagreened, with golden-green to purple-blue sheen ( Figs. 364 View FIGURES 362 – 366 , 399 View FIGURES 395 – 400 ), or micropapillose, almost matt, with golden or greenish metallic tinge ( Figs. 181 View FIGURES 181 – 186 , 193 View FIGURES 187 – 193 , 245, 251 View FIGURES 244 – 252 , 261 View FIGURES 253 – 261 , 276 View FIGURES 276 – 281 , 291 View FIGURES 282 – 291 ). Male abdomen with 5, female with 6 visible segments. Male sternites non-modified, subrectangular, slightly transverse, sternite 5 trapezoid ( Figs. 274 View FIGURES 268 – 275 , 359 View FIGURES 354 – 361 ).
Female tergite 2 laterally with pair of dimple-like structures lacking metallic sheen and having hexagonal cellular structure ( Figs. 85 View FIGURES 80 – 88 , 120 View FIGURES 114 – 123 , 163 View FIGURES 158 – 166 , 290 View FIGURES 282 – 291 , 333 View FIGURES 328 – 336 ); female sternites non-modified, subrectagular; sternites 3–6 without anteromedial apodemes. Female tergite and sternite 6 short and transverse, subequal, conspicuously narrower and shorter than tergite 5.
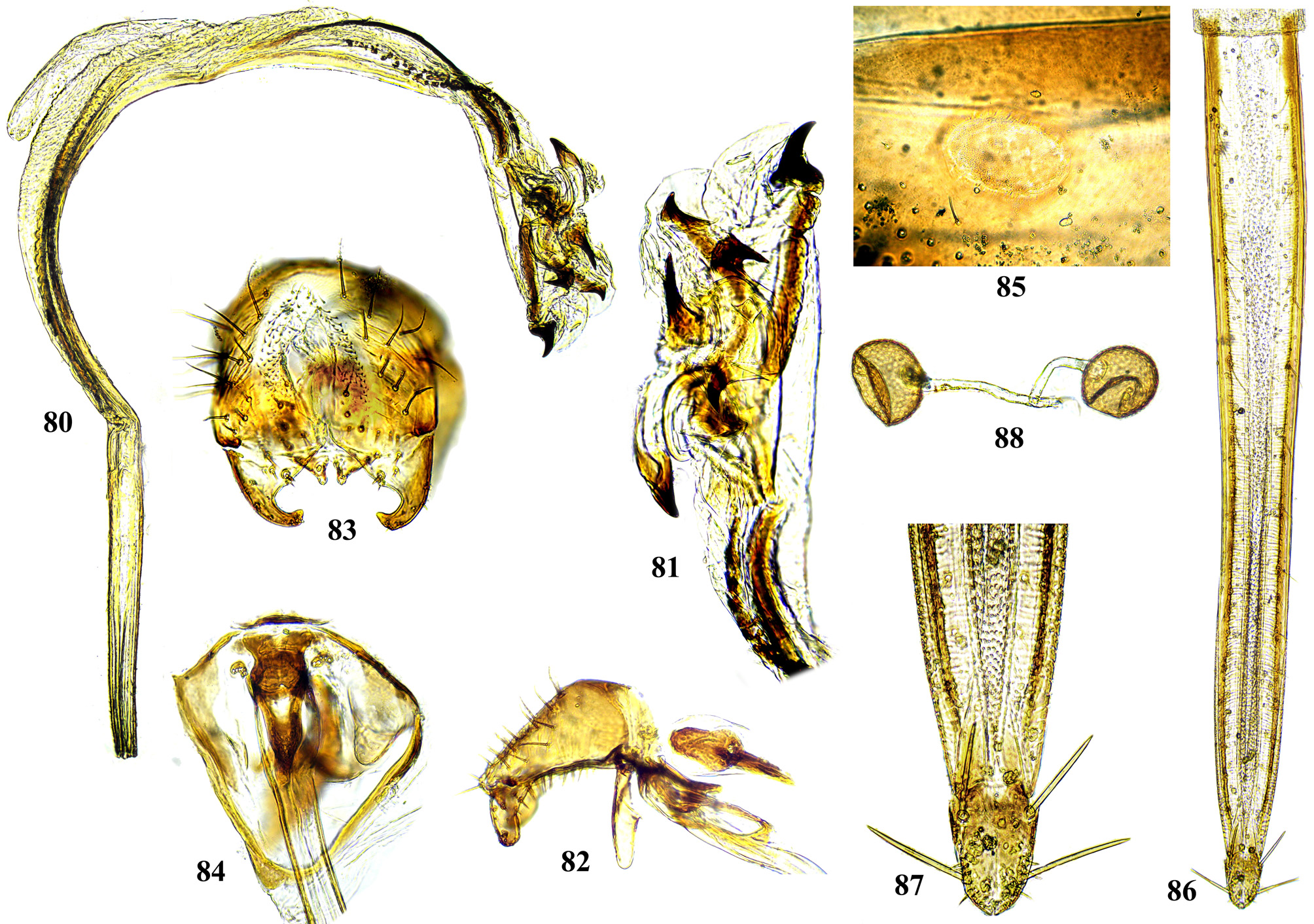
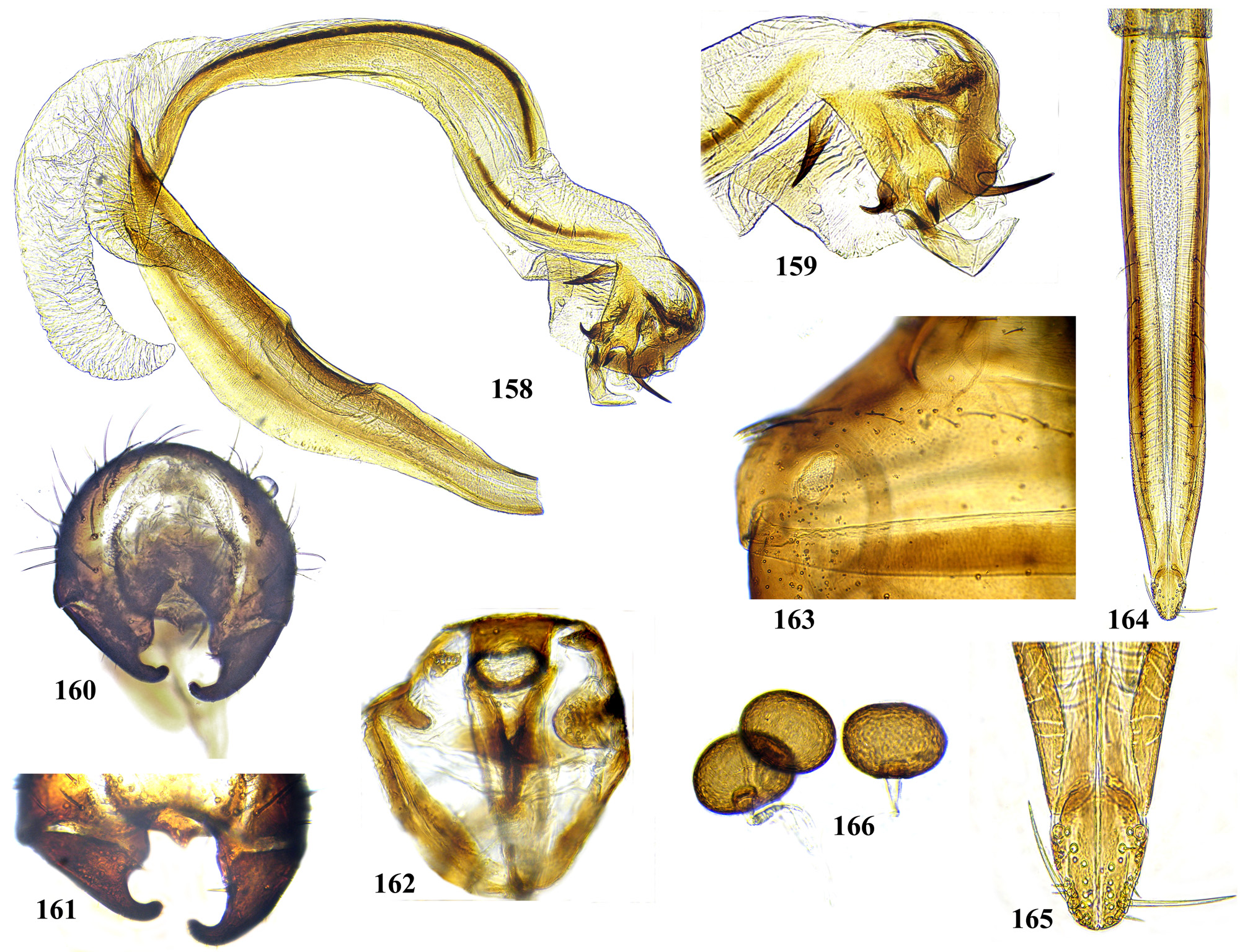
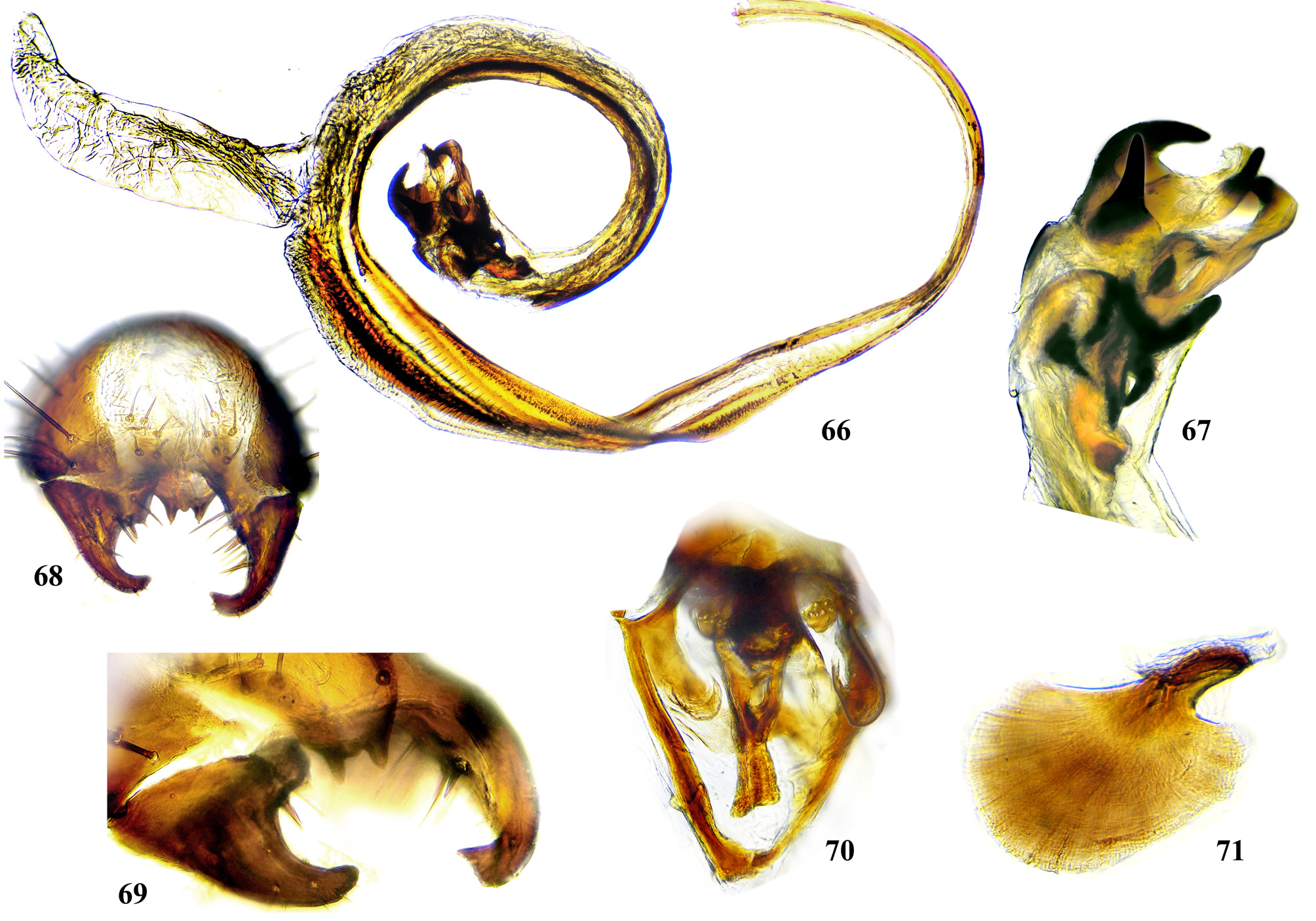
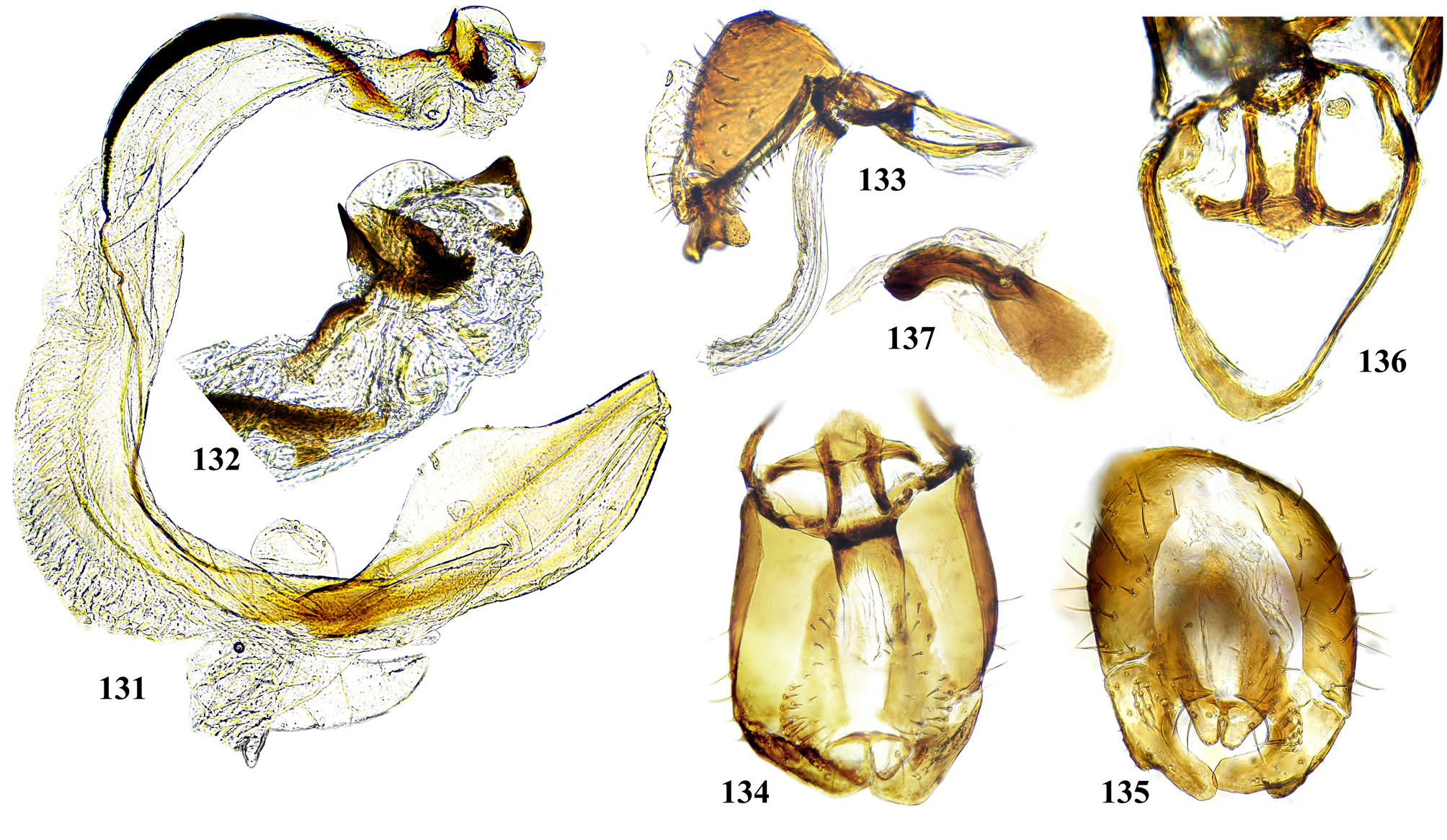
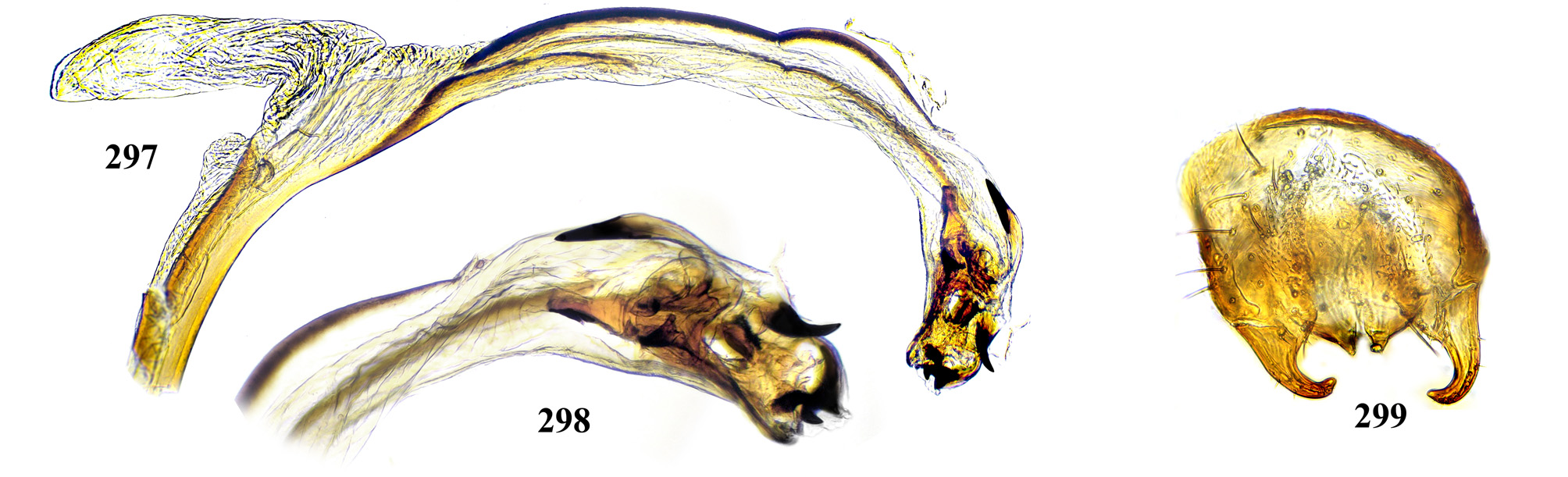
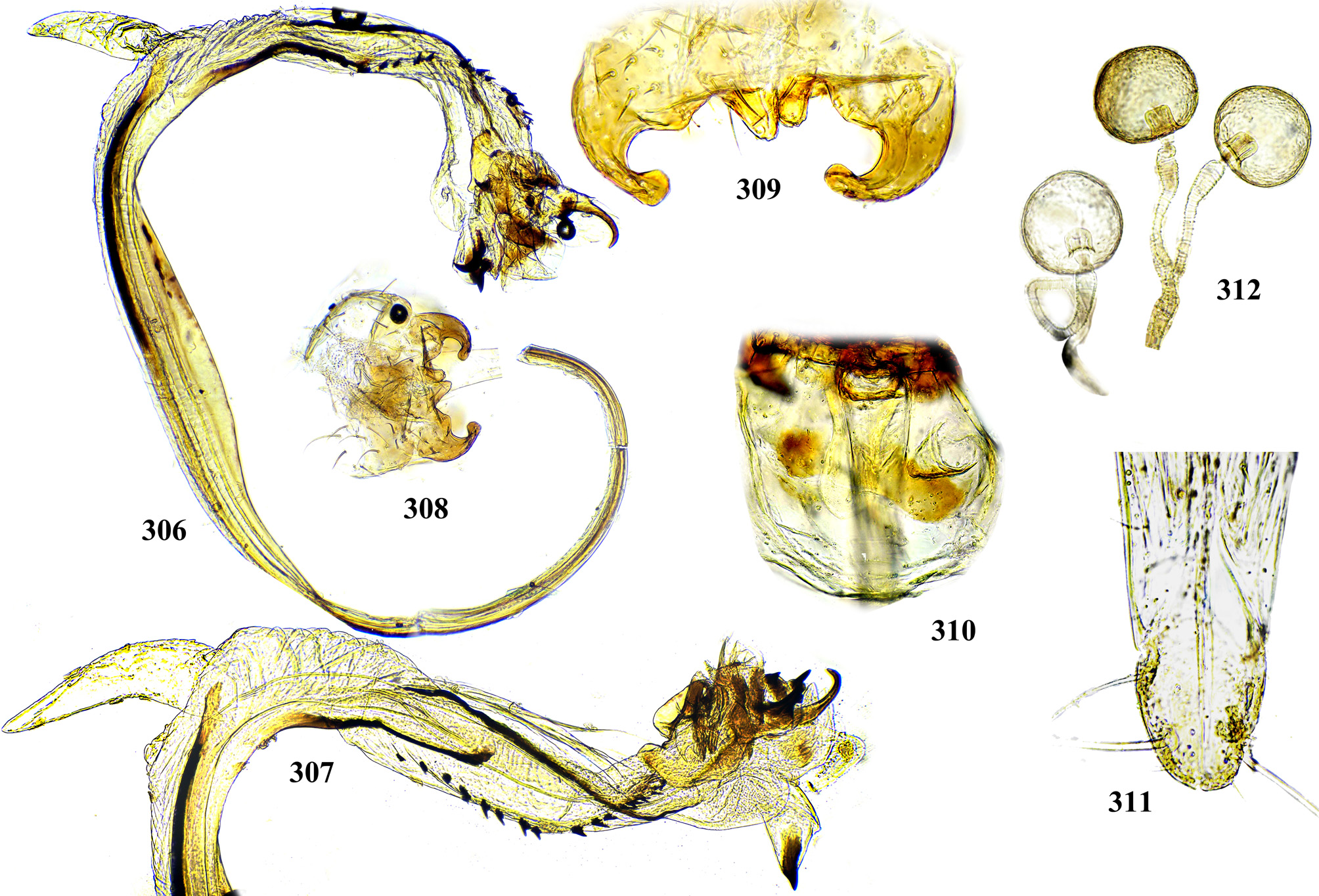
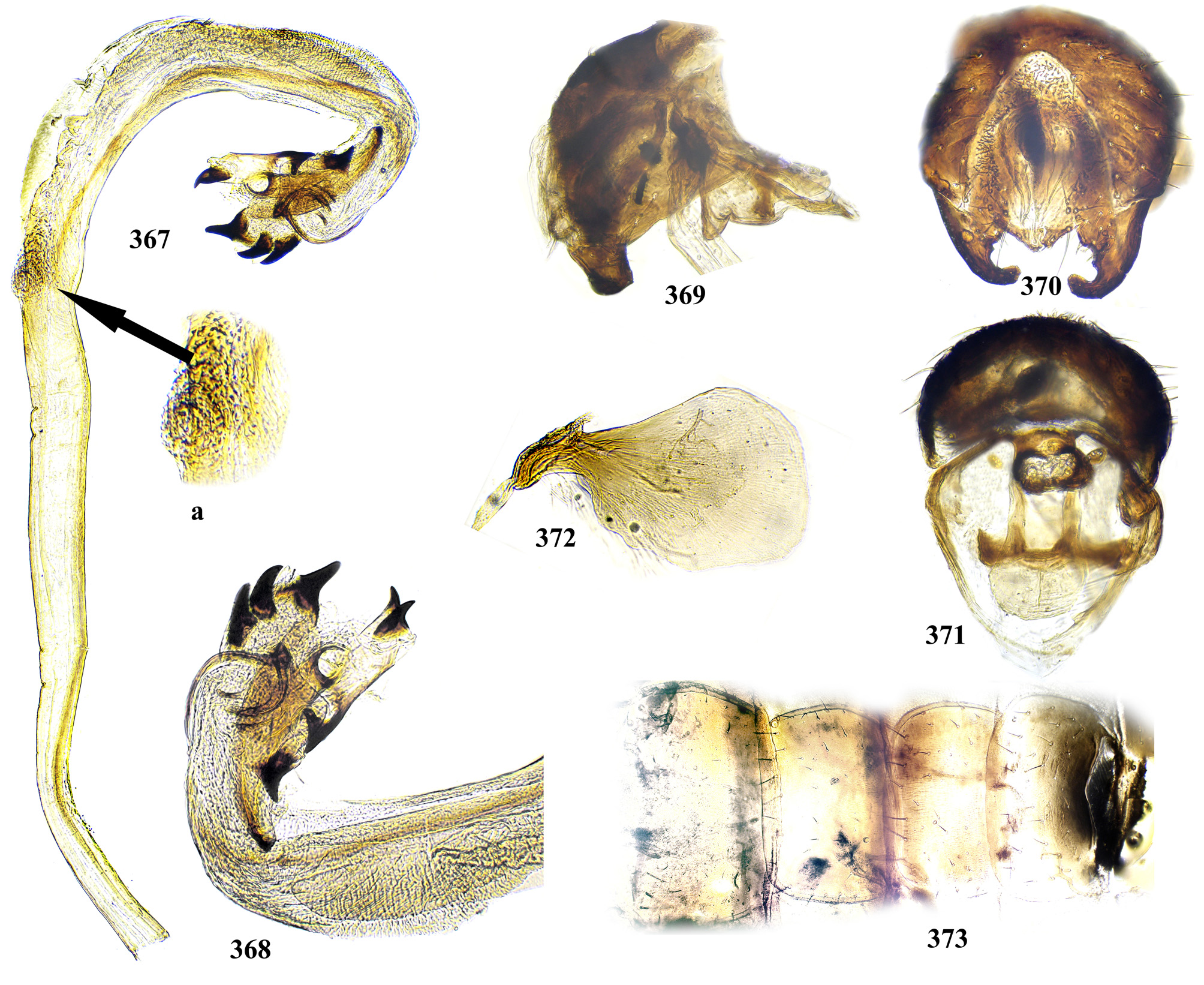
Male postabdomen. Membranous pouch between tergite 5 and postabdomen, dorsolateral, densely microtrichose ( Fig. 225 View FIGURES 223 – 230 ); sternite 8 setulose ( Fig. 260 View FIGURES 253 – 261 ). Epandrium small. Surstyli short, separated from epandrium by seam, wide basally, strongly narrowed and curved mesally towards apex ( Figs. 48 View FIGURES 46 – 52 , 68 View FIGURES 66 – 71 , 83 View FIGURES 80 – 88 , 100 View FIGURES 97 – 105 , 117 View FIGURES 114 – 123 , 161 View FIGURES 158 – 166 , 176 View FIGURES 174 – 180 , 210 View FIGURES 210 – 216 ), usually pointed or serrate apically ( Figs. 191 View FIGURES 187 – 193 , 212 View FIGURES 210 – 216 ), in P. euphorbiana Krivosheina & Krivosheina with pair of blunt lobes ( Figs. 133, 135 View FIGURES 131 – 137 ); usually without basimesall lobe, only in P. longicornis Hendel such lobe present ( Fig. 226 View FIGURES 223 – 230 ). Proctiger low, not protruding posteriorly, usually with pair of short submedial nipple-like ventral projections ( Figs. 33 View FIGURES 31 – 40 , 48 View FIGURES 46 – 52 , 68 View FIGURES 66 – 71 , 83 View FIGURES 80 – 88 , 135 View FIGURES 131 – 137 , 177 View FIGURES 174 – 180 , 190 View FIGURES 187 – 193 , 202 View FIGURES 200 – 205 , 256 View FIGURES 253 – 261 , 271 View FIGURES 268 – 275 , 299 View FIGURES 297 – 299 , 309 View FIGURES 306 – 312 , 349 View FIGURES 346 – 349 ) bearing single apical short setula ( Fig. 33 View FIGURES 31 – 40 ) (absent in P. leucotricha Kameneva & Korneyev ); nipple-like processes fused anteriorly in P. longicornis ( Figs. 226–228 View FIGURES 223 – 230 ); or strongly enlarged and fused in P. euphorbiana ( Fig. 135 View FIGURES 131 – 137 ) cerci flat and not fused in P. spriggsi sp. n. ( Fig. 356 View FIGURES 354 – 361 ). Subepandrial sclerite flat, V- or U-shaped ( Figs. 34 View FIGURES 31 – 40 , 100 View FIGURES 97 – 105 , 134 View FIGURES 131 – 137 , 179 View FIGURES 174 – 180 , 203 View FIGURES 200 – 205 , 227 View FIGURES 223 – 230 ), without thickened tooth-like setulae (prensisetae), with at most one thickened seta inserted into larger alveola ( Figs. 48 View FIGURES 46 – 52 , 83 View FIGURES 80 – 88 , 177 View FIGURES 174 – 180 , 202 View FIGURES 200 – 205 ). Hypandrium posteriorly fused or joined by membrane to sclerotized arms of epiphallus; U-shaped, usually asymmetrical: right gonite ovoid, devoided of setulae, with sclerotized lateral sclerite, left gonite inconspicuous; phallapodeme usually Y-shaped, vanes inconspicuous, at right side represented by sclerotized bar, and entirely reduced at left side; in P. euphorbiana , P. spriggsi sp. n. and P. steyskali sp. n. arms of phallapodeme widely spaced, vanes almost symmetrical, and phallapodeme itself flattened antero-posteriorly rather than laterally ( Figs. 136 View FIGURES 131 – 137 , 358 View FIGURES 354 – 361 , 371 View FIGURES 367 – 373 ). Sensillar plates at sides of phallus base rounded, simple.
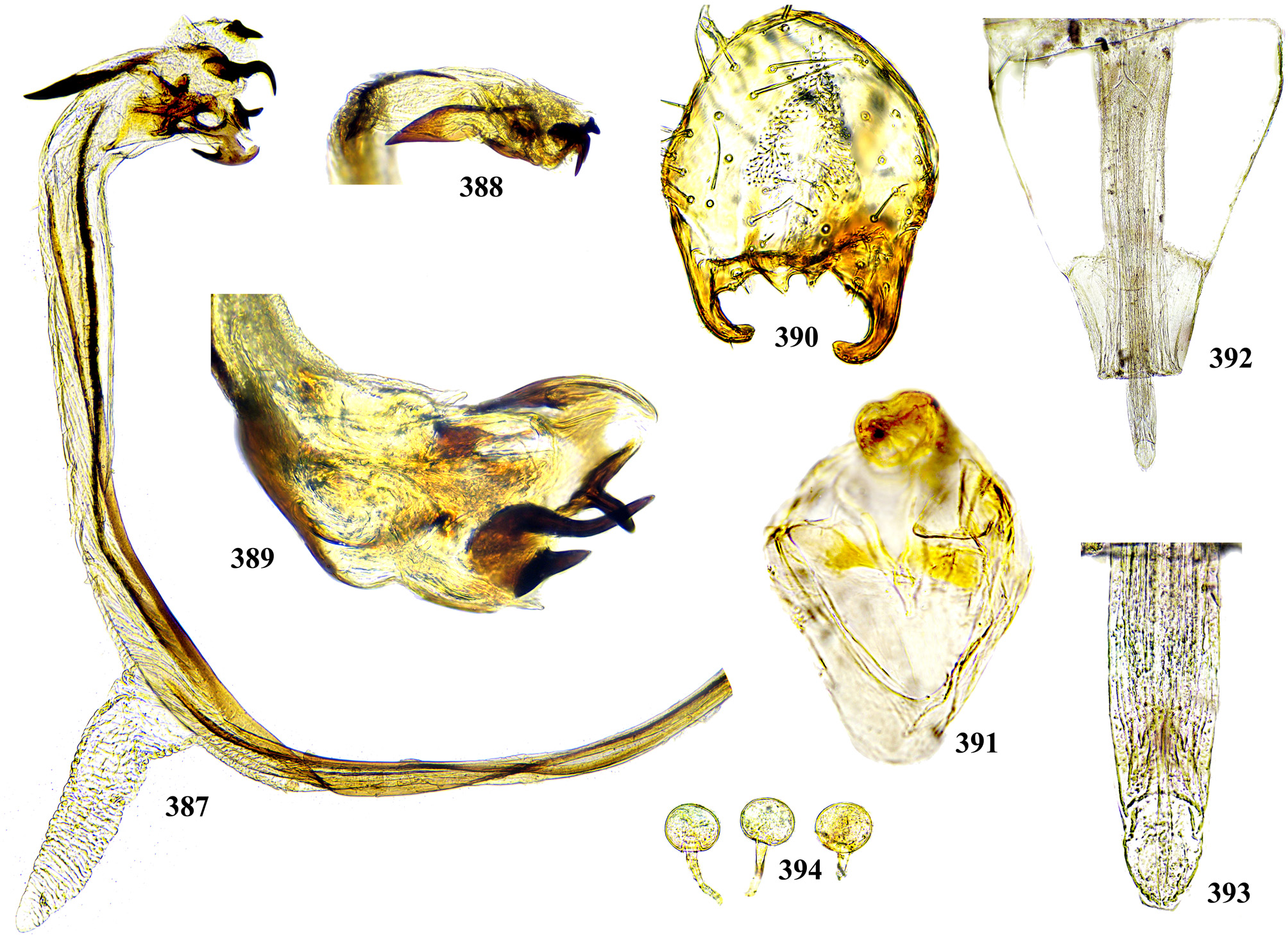
Phallus is the stucture strongly involved into diagnostics of species in the tribe Ulidiini, and detailed description of its structures, which are rather different from those in other Tephritoidea, is provided here. It is a very large tubular organ, usually 1.2–1.5 times longer than male abdomen, with thick ring of basiphallus joined by two sclerotized lateral arms (derivatives of the epiphallus?) to the postero-medial branches of hypandrium. Remaining part of the phallus represented by distiphallus is a bare flattened tube with a pair of sclerotized taeniae, one reaching approximately the middle of phallus length, and another running throughout almost whole length of the phallus. The blind memranous projection, or caecum, situated at middle of phallus length and being everted by high pressure of haemolymph before whole phallus itself is erected, believed to serve for its penetration into female aculeus and vagina, is known exclusively in Ulidiini; caecum of Physiphora always single, long ( Figs. 66 View FIGURES 66 – 71 , 97 View FIGURES 97 – 105 , 158 View FIGURES 158 – 166 , 223 View FIGURES 223 – 230 , 237 View FIGURES 237 – 243 , 253 View FIGURES 253 – 261 , 346 View FIGURES 346 – 349 ), short ( Figs. 31 View FIGURES 31 – 40 , 46 View FIGURES 46 – 52 , 306 View FIGURES 306 – 312 , 328 View FIGURES 328 – 336 , 354 View FIGURES 354 – 361 ) or vestigial ( Fig. 367 View FIGURES 367 – 373 a) (similar structures are known in both Tephritidae and Platystomatidae , but are located subapically, on preglans or glans). The part of distiphallus basal of the caecum is called stipe (the term was applied by McAlpine (1973) to the homologous part of the Platystomatidae phallus). The part of distiphallus apically of the caecum consists of the preglans and glans. The preglans is a tubular section of the distiphallus bearing the continuation of one basal taenia not reaching the glans, and the separate short preglandic taenia usually joined to the glans or forming its body. The membrane of preglans is usually strongly wrinkled and often bearing tiny or strong spines ( Figs. 31 View FIGURES 31 – 40 , 46 View FIGURES 46 – 52 , 80 View FIGURES 80 – 88 , 175 View FIGURES 174 – 180 , 224 View FIGURES 223 – 230 , 268 View FIGURES 268 – 275 , 306 View FIGURES 306 – 312 ). The preglans of Physiphora is shorter ( Figs. 31 View FIGURES 31 – 40 , 66 View FIGURES 66 – 71 , 80 View FIGURES 80 – 88 , 97 View FIGURES 97 – 105 , 200 View FIGURES 200 – 205 , 223 View FIGURES 223 – 230 , 268 View FIGURES 268 – 275 , 306 View FIGURES 306 – 312 , 367 View FIGURES 367 – 373 ) or longer ( Figs. 46 View FIGURES 46 – 52 , 131 View FIGURES 131 – 137 , 297 View FIGURES 297 – 299 , 387 View FIGURES 387 – 394 ) than the stipe, or sometimes they are of subequal length; this character is important for identification of some species. Glans is the apical structure represented by sclerotized spines, hooks, flaps, petals, and lobes of different shape surrounding the gonopore. The glans can be formed mainly by the expanded apex of taenia and a few short spines, as in P. elbae Hendel, P. leucotricha Kameneva & Korneyev, and P. hendeli sp. n. ( Figs. 116 View FIGURES 114 – 123 , 175 View FIGURES 174 – 180 , 215 View FIGURES 210 – 216 ) or a separate inner sclerites and some prominent but rather short lobes as in P. euphorbiana Krivosheina & Krivosheina and P. maraisi sp. n. ( Figs. 131 View FIGURES 131 – 137 , 238 View FIGURES 237 – 243 ), or mostly by numerous long or very long acute spines, as in P. kirki sp. n., P. meyi sp. n., P. opalizana sp. n., P. sericea Hendel , and P. smaragdina Loew ( Figs. 201 View FIGURES 200 – 205 , 254 View FIGURES 253 – 261 , 283 View FIGURES 282 – 291 , 328–329 View FIGURES 328 – 336 , 346– 348 View FIGURES 346 – 349 ).
Ejaculatory apodeme usually wide fan-like, with short apical portion and relatively small sperm pump ( Figs. 71 View FIGURES 66 – 71 , 119 View FIGURES 114 – 123 , 205 View FIGURES 200 – 205 , 230 View FIGURES 223 – 230 , 243 View FIGURES 237 – 243 , 275 View FIGURES 268 – 275 , 332 View FIGURES 328 – 336 , 372 View FIGURES 367 – 373 ).
Female oviscape short, at most as long as tergite 5 and 0.25–0.3 times as long as aculeus. Eversible membrane ( Fig. 35 View FIGURES 31 – 40 , 50 View FIGURES 46 – 52 ) as long as aculeus, with 2 pairs of bare taeniae almost reaching its posterior margin; membrane finely microtrichose in anterior 0.8 and covered with fine multidentate scales in posterior 0.2 of its length. Aculeus ( Figs. 36 View FIGURES 31 – 40 , 51 View FIGURES 46 – 52 , 86 View FIGURES 80 – 88 , 102 View FIGURES 97 – 105 , 121 View FIGURES 114 – 123 , 164 View FIGURES 158 – 166 , 287 View FIGURES 282 – 291 , 334 View FIGURES 328 – 336 , 360 View FIGURES 354 – 361 ) narrow and moderately long, 6.5–12 times as long as wide, with elongate oval cercal unit bearing 2 pairs of long setae (basal dorsal and subapical lateroventral) and several shorter setulae on ventral and latero-ventral side; anal opening (slit) on dorsal side of cerci ( Figs. 37 View FIGURES 31 – 40 , 87 View FIGURES 80 – 88 , 103 View FIGURES 97 – 105 , 165 View FIGURES 158 – 166 , 288 View FIGURES 282 – 291 , 311 View FIGURES 306 – 312 , 335 View FIGURES 328 – 336 ); sternite 8 well-sclerotized, with 11–12 setulae on each side. Medial groove covered with monodentate scales.
Vagina with simple, finger-like ventral receptacle ( Figs. 38–39 View FIGURES 31 – 40 , 104 View FIGURES 97 – 105 ). Three spherical (or collapsed to mushroom-like shape) spermathecae, single right and 2 left spermathecae on Y-shape bifurcated common duct ( Figs. 40 View FIGURES 31 – 40 , 52 View FIGURES 46 – 52 , 105 View FIGURES 97 – 105 , 122 View FIGURES 114 – 123 , 166 View FIGURES 158 – 166 , 289 View FIGURES 282 – 291 , 312 View FIGURES 306 – 312 , 336 View FIGURES 328 – 336 , 361 View FIGURES 354 – 361 , 394 View FIGURES 387 – 394 ); short, invaginated “necks” at junction of spermatheca with ducts.
Preimaginal stages. Morphological details of puparium of P. alceae Preyssler were originally figured by Hennig (1940: Textfig. 8–10, as P. demandata ). Later, Krivosheina & Krivosheina (1997: 460, Figs. 4 View FIGURES 1 – 4 –5) described the third instar larva and puparium of P. chalybea Hendel.
Egg of P. elbae ( Fig. 123 View FIGURES 114 – 123 ) elongate-ovoid, 0.65–0.67 mm long, 4.0–4.2 times as ling as wide, bluntly rounded at both apices, without visible sculpture; similarly, egg of P. alceae of the same shape, 0.68–0.78 mm, slightly arcuate dorsally, with micropile shielded dorsally by tubercle (Allen & Foote 1967).
Third instar larva of P. alceae 4.64–7.7 mm long (Allen & Foote 1967), in P. chalybea (after Krivosheina & Krivosheina 1997) white, narrow, 7 times as long as wide, bluntly truncated at posterior end; gnathocephalon with numerous deep and straight oral ridges; sensory organs not described; cephalopharyngeal skeleton with mandibular sclerite almost four times as long as high, with single very long and blunt mouthhook and without additional lobes or teeth; hypostomal (= hypopharyngeal) sclerite long, six to seven times times as long as high, parastomal sclerite as long as hypostomal sclerite, entirely separated from it; dental sclerite present, straight and slender; labial (= subhypostomal) sclerite very long, bare-like in profile; pharyngeal cornua poorly sclerotized; cephalopharyngeal skeleton of P. alceae of same shape, showing no essential differences (Hennig, 1940: Textfig. 8; Allen & Foote 1967: Figs. 17–19, as P. demandata ). Anterior spiracle of P. chalybea with ten lobes (papillae) (Krivosheina & Krivosheina 1997: Figs. 4.2 & 5.7); in P. alceae with 12–14 lobes (Allen & Foote 1967). Abdominal segments with creeping welts covered by denticles (Krivosheina & Krivosheina 1997: Fig. 4.2). Anal plate of P. alceae broadly triangular, bordered by 2–3 rows of spinules (Allen & Foote 1967). Posterior spiracular plates similar in P. alceae and P. chalybea , not raised, with subparallel, oblique spiracular slits (inner slit at 30º to dorso-ventral axis. Intraspiracular hairs (four) triple, ramulose at apices.
Puparium orange, strongly wrinkled (Krivosheina & Krivosheina 1997: Fig. 5).
Larval feeding remains poorly known even in P. alceae , the best examined species. Despite early reports of larval feeding in corn silage (Drake & Decker 1932) and rotting narcissus bulbs (Blanton 1938), further laboratory experiments showed that larval feeding failed in rotting plant matter alone, but was successful in dung or dung mixed with vegetation; larval growth lasted 25–30 days, of them 3rd instar larvae lived c. 10 days; prepupal stage lasts one day, and the pupal period is 15 days (Allen & Foote 1967). P. clausa was reared from larva feeding in compost (see the labels below).
Larvae of P. chalybea were found under the bark of the Desert Poplar, ( Populus euphratica Oliv. ). Overwintered larvae form clusters under the bark of a fallen tree. Larvae are apparently saprophagous, casually eating dead beetle larvae (Krivosheina & Krivosheina 1997).
In addition, adults of P. euphorbiana Krivosheina & Krivosheina were reared from rotting tissues of the succulent shrub Euphorbia royleana Boiss. in India and Pakistan (see below).
The label data accumulated in this study clearly show that larvae of several paleotropical species, e.g., P. allomma Speiser, P. clausa Macquart, P. longicornis Hendel , and P. violacea Hendel , infest growing tips or feed in the rotting stems of palms, mainly of the date-palm. In addition, even P. alceae known to feed as larva mostly in compost or dung, has been reared in Saudi Arabia from rotting date-palm, the typical substrate of many other Physiphora . Such mode of larval feeding, possibly in assiciation with the burrow froth and feces of the palm weevils Rhynchophorus ferrugineus (Olivier 1790) is believed to be common for most widespread Physiphora species.
Adult flies are usually attracted to bird droppings, dung or human feces, and in some cases can be captured in Malaise or light traps.
Mating behaviour of P. alceae was shown to include moving by straightened fore and mid legs by the male, vibration by the wings, dancing before female, obviously to demonstrate the face and eye pattern as well as colours of body sheen to her. Final part of courtship includes pushing female face by males abdomen, followed by sucking of liquid from male’s posterior end1 (Alcock & Pyle 1979).
Phylogenetic relationships. Physiphora possesses at least two unique possible synapomorphies that support its monophyly: the nipple-like process on cerci of male and the dimple-like structure on tergite 2 of females; other synapomorphies being subject to homoplasy (parafrontal microtrichose areas, scutum and abdominal tergites mostly devoided of setae) also support its monophyly, whereas the presence of deeply impressed and ventrolaterally delimited antennal grooves possibly support monophyly of Ulidia + Timia , the largest lineage in the tribe Ulidiini.
The relationships within the genus hardly can be inferred from the morphological characters, concerning that their number (10–12 characters for almost 30 species) is far from sufficient. At most, 3 groups can be intuitively selected as possibly monophyletic: clausa + flavipes + violacea (cell r4+5 closed, and petiolate—unique synapomorphy), euphorbiana + steyskali (frons widely microtrichose,—rare character, also only in P. leucorticha (possibly due to homoplasy) and phallapodeme with parallel arms and well developed and symmetrical vanesunique synapomorphy), and hendeli + obscura (phallus with finely spinulose preglans—unique synapomorphy).
Geographical distrbution. Physiphora alceae and P. clausa are widespread in almost all zoogeographical regions. There are 18 species of Physiphora occurring only in the mainland Subsaharan Africa. Two species, P. azurea and P. tarsata , are endemics of the Seyshelles and Reunion. Only P. chalybea Hendel is rescticted to Central Asia. Physiphora aperta Steyskal is endemic to Solomon Islands. Five species were listed from the Oriental Region, one of them ( P. longicornis Hendel ) occurs from Taiwan and Ceylon, and P. euphorbiana Krivosheina & Krivosheina was found in Indian Subcontinent. P. sericea is distributed from Eastern Africa through Arabian Peninsula to Thailand. P. leucotricha is known by far only from the Arabian Peninsula, whereas P. smaragdina is widespread in the whole Africa, Near East, and Arabian Peninsula. Finally, an apparently undescribed species closely related to P. violacea (P. sp. A), has been recently recorded from Australia, but additional material is needed to see if it is a local endemic or another adventive species.
Taxonomic and nomenclatural remarks. The genus was partly revised by Hendel (1909; 1913b); later, Séguy (1941) described and keyed several Afrotropical species. Recently, Chen & Kameneva (2007) revised Chinese species, and Kameneva & Korneyev (2010) gave a review of the species from the United Arab Emirates. However, Afrotropical fauna needed more thorough treatment, with the use of the phallus characters, which were found to be constant and important for identification of species.













































No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ulidiinae |
|
Tribe |
Ulidiini |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ulidiinae |
|
Tribe |
Ulidiini |