
Sturisoma barbatum ( Kner, 1853 )
|
publication ID |
https://doi.org/10.11646/zootaxa.5360.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:3B49D165-F9D5-4697-A344-D4D7977F60A3 |
|
DOI |
https://doi.org/10.5281/zenodo.10164909 |
|
persistent identifier |
https://treatment.plazi.org/id/52178793-D800-FFB4-01DB-C9D0FBC2F962 |
|
treatment provided by |
Plazi |
|
scientific name |
Sturisoma barbatum ( Kner, 1853 ) |
| status |
|
Sturisoma barbatum ( Kner, 1853) View in CoL
Original description. Loricaria barbata Kner, 1853: 12 View in CoL , 23, pl. 5.
Type locality: Rio Cuiabá , Cujaba-Fluss [ Brazil]. Holotype: NMW 46155 ( Fig. 1 A–B View FIGURE 1 and Fig. 2 View FIGURE 2 )
Oxyloricaria robusta ( Regan, 1904) View in CoL : 300, pl. 19 (fig. 1).
Type locality: Paraguay river . Lectotype: BMNH 1895.5.17.89 and Paralectotype BMNH 1895.5.17.90 ( Fig. 1 C View FIGURE 1 and Figure 3 A–B View FIGURE 3 )
Sturisoma robustum ( Eigenmann, 1907) View in CoL , Sturisoma robustum ( Eigenmann, 1909) View in CoL , Oxyloricaria robusta Pozzi, 1945 View in CoL , Oxyloricaria barbata Pozzi, 1945 , Sturisoma robusta ( Arámburu et al., 1962) View in CoL , Sturisoma robusta Ringuelet et al., 1967 View in CoL .
Examined material ( Fig. 4 View FIGURE 4 ). ARGENTINA: Salta: CFA-IC-5103 (1) Pilcomayo Basin, in Santa Victoria close to La Merced. Col. Yamila Cardoso, Sergio Bogan, and Juan M. Meluso (FHN-2416) 22°13’28.70”S / 62°40’48.94”W, 16/10/2015 GoogleMaps ; CFA-IC-5202 (1) Pilcomayo Basin in Santa María . Col. Yamila Cardoso, Sergio Bogan, and Juan M. Meluso (FHN-2379), 22° 8’7.73”S / 62°48’45.18”W), 16/10/2015; CFA-IC-5253 (8) Bermejo Basin in La Cañada Stream and Ruta Provincial 15. Col. Yamila Cardoso, Sergio Bogan, and Juan M. Meluso, (FHN-2583 a 2590), 23°21’6.29”S / 64° 1’16.32”W, 17/10/2015 GoogleMaps . Formosa: CFA-IC-3133 (2), Paraguay Basin in Formosa City. Localidad 9. Col. Y. Cardoso, A. Paracampo, C. Rivera & J. Montoya-Burgos (AR11-604 and 605) 26° 12, 016’ S / 58° 08,776’ W, 25/11/2011 GoogleMaps ; CFA-IC-3396 (2) Bermejo Basin . Locality 7. Col. Y. Cardoso, A. Paracampo, C. Rivera & J. Montoya-Burgos (AR11-326), 26° 20, 217’ S / 59° 21,170’ W, 22–23/11/2011 GoogleMaps ; CFA-IC-6380 (1) Pilcomayo Basin , Canal of Pilcomayo River close to El Quebrachal. Punto 24. Col. Sergio Bogan and Juan M. Meluso, (FHN-3318), 23°18,529´S / 61°50,576´W, 19/05/2016 GoogleMaps . CFA-IC-6381 (1) Pilcomayo Basin , Canal of the Pilcomayo River close to El Quebrachal. Punto 23. Col. Sergio Bogan and Juan M. Meluso (FHN-3298), 23°19,195´S / 61°50,118´W, 19/05/2016 GoogleMaps ; CFA-IC-8731 (1) Pilcomayo Basin , en cruce Canal, up to Bañado La Estrella. Col. A. Paracampo and D. Barrasso, 25° 8’ 24,4’’ S / 61° 57’ 7’’ W, 25/08/2006 GoogleMaps . Santa Fe: CFA-IC-6268 (1) Paraná Basin , Colastine River . Col. Sergio Bogan, 31° 39.686’S / 60° 35.767’W, agosto de 2007; GoogleMaps MACN-IC-7739 (1), Colastiné River , 1993; GoogleMaps MLP-9632 (4) Colastiné River ; GoogleMaps MLP-9631 (2) Coronda River , Dto Capital. Col. R. Peruso; GoogleMaps MLP-9180 (1) Rosario. Col. C. Vidal, 1960 GoogleMaps . Corrientes: MACN-IC-7282 (1) Corza Cué, Paraná River . Col. J.O. Fernández Santos, 27/07/1927 ; MACN-IC-7282 (7) Corza Cué, Paraná River . Col. J.O. Fernández Santos, 20/07/1927 . Chaco: MLP-8088 (1) Laguna Grande , Resistencia. Col. M. Galvan, 2/12/1966 ; MLP-8105 (2) Balneario Rio Negro , Resistencia. Col. M. Galvan, 12/12/1966 ; MLP-8969 (1) Paraná River ; MACN-IC-7282 (1) MACN-IC-677 (4) Paraguay River . Col. Zotta, Migoya and Palavecino, September 1929 . PARAGUAY: BMNH 1934.8.20.389-391 , Paraguay River , close to Asunción, Col. Dr G. B. Schouten, 1934.
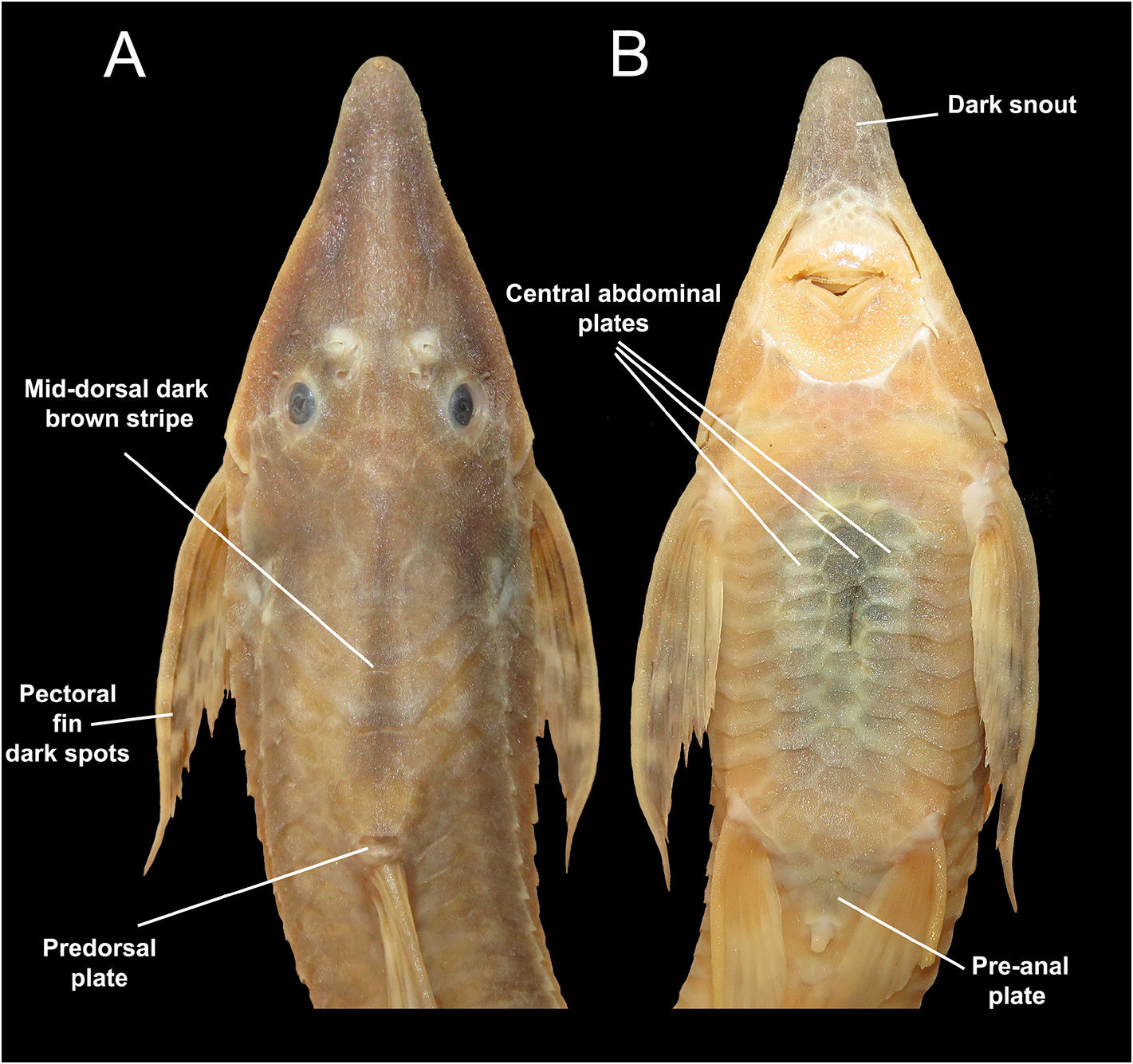
Emended diagnosis. Sturisoma barbatum is diagnosed by the following combination of characters: 32–33 median plates and a supracaudal plate; first 17–19 (most frequently 17) mid-series plates ( vs. 21–22 in S. rostratum and S. monopelte ); 14–15 plates with fused keels ( vs. 16–17 in S. tenuirostre ); 18–19 ventromedial lateral plates; seven to nine lateral abdominal plates; anteriormost abdominal plates, just posterior to the lower lip, square, not enlarged, smaller than anterior abdominal plates (shared with S. reisi vs. rectangular, enlarged, longer than anterior abdominal plates); absence of ventral (see Ghazzi 2005) process in the sphenotic (shared with S. tenuirostre and S. graffini vs. well-developed process in S. guentheri , S. nigrirostrum , S. monopelte , S. rostratum , S. lyra ); premaxillary teeth subequal or smaller than dentary teeth ( vs. premaxillary teeth larger in S. graffini ); frontal bone contributing less than half the length of the dorsal margin of the orbital crest (shared with S. graffini and S. guentheri , vs. frontal more than half the length of the dorsal margin of the orbital crest in remaining species); presence of dark spot in the first three radii of the pectoral and the first two radii of the anal fins (shared with S. graffini and S. reisi ).
Description
Long, narrow body, completely covered by dermal plates ( Fig. 6 View FIGURE 6 ). Greatest height of body at origin of dorsal fin. Depressed caudal peduncle. Ventral margin in lateral view flat to slightly convex at beginning of caudal section. Subtriangular head with broad, rounded snout. 32–33 median plates (17–19 without fused keels, remaining 15 with fused keels) and supracaudal long, triangular, and extending over median rays of caudal fin. Seven to nine (median eight) lateral abdominal plates angulated between last ray of pectoral fin and origin of pelvic fin. Anteriormost abdominal plates, just posterior to lower lip, square, not enlarged, smaller than anterior abdominal plates. Three central series of relatively large abdominal plates, quadrangular to subcircular in contour, arranged in well-defined rows. Preanal plate bordered anteriorly by three well-developed plates ( Fig. 6 View FIGURE 6 ).
Orbit subcircular without postorbital notch. Sphenotic without lateral process. Nostrils positioned dorsally and located in groove, their anterior part forms a tubule of skin that projects dorsally and whose posterior edge is higher, forming a small shelf of skin. Surface of the lip covered by round papillae. Corner with a small maxillary barbel, which generally does not exceed the diameter of the orbit. Slender, bifid teeth, 30–38 (mean 36) in the premaxilla, 27–33 (mean 29) in the dentary, visibly curved cusps, larger lateral cusp, and slightly rounded margins. The teeth of the premaxilla and dentary of the same size. The largest symphyseal teeth in dentary ( Fig. 6 View FIGURE 6 ).
Origin of dorsal fin at the same level or slightly anterior of pelvic origin fin. Dorsal fin with seven branched rays, and a relatively long and thin spine that continues into a rather short filamentPectoral fins with narrow and long first ray as filament ( Fig. 8B View FIGURE 8 ) and six branched, gently curved rays. Pectoral fin reaches or exceeds the origin of pelvic fin. Pelvic fin long, with first ray longest, and widely extending beyond the origin of anal fin, has an unbranched first ray followed by five branched rays. Caudal fin forked, with concave posterior edge and both upper and lower lobes continuous in long and delicate filaments, with 12 branched rays.
Ventrally, pale yellow to orange body, especially on the abdomen. In some specimens, abdomen shows faded dark spots. Ventral surface of snout with conspicuous dark pigmentation ( Fig. 5 View FIGURE 5 ).
Dorsal and pectoral fins with light brown membranes, irregular brown spots on the membrane covering spine and first branched rays; other rays may also have light brown spots. Pelvic and anal rays with light-toned membranes and brown spots or bands in first rays. Caudal fin with dark band onto the upper and lower external rays, interrupted by light spots (see also Almirón et al., 2015; Monasterio de Gonzo 2003). The fixed specimens have a variable background color between yellowish, ocher, and brown, depending on state of conservation. Back is generally darker. Dorsolateral strip in some cases well-defined and in others diffuse. Caudal peduncle with the interruption of the dorsolateral dark stripes noted in posterior end of the keels. Compound pterotic, a high concentration of melanophores. The opercular area lighter.
Sexual dimorphism. Males with well-developed, long, slender odontodes that project backward along sides of head; longest odontode just anterior to cleithrum ( Fig. 7 View FIGURE 7 and 8 View FIGURE 8 ). Kner (1853: 24) described this as a thick beard with whitish bristles of unequal length curved backward; these bristles sink inserted into odontode sheats that do not reach below the lateral edge of the head. Pectoral spines also with hypertrophied odontodes shorter than those of head ( Fig. 8 View FIGURE 8 ). Males with greater development of odontodes that form median keels, especially notable in coalescing plates.
Distribution
Sturisoma barbatum is distributed in the Paraguay River Basin in Brazil, the Republic of Paraguay, and Argentina. Particularly, in Argentina, it occurs in the Pilcomayo, Bermejo, and Paraguay basins in Salta, Formosa, and Chaco provinces (also see Liotta 2005; Monasterio de Gonzo 2003). In the Middle and Lower Paraná River, this species was recorded from Misiones, Corrientes, Entre Ríos ( Almirón et al., 2015), and Santa Fe provinces. Its southernmost occurrence was published by Ringuelet et al. (1967) from the city of Rosario.
Discussion
As follows, we checked all previously conducted studies, specimens used, and taxonomic decisions taken throughout the taxonomic history of S. barbatum .
The original description of S. barbatum Kner (1853) was based on a single male specimen of 244 mm, with highly developed odontodes on the sides of the head (also noticeable on the pectoral spines), traits that were included in the diagnosis ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ). The description has morphological features that Kner used to distinguish between S. barbatum and S. rostratum . Kner (1853: 23) indicated that these two species share many features of the plates of the lateral series: “both lateral keels acute, finely serrated, especially towards the tail where the last 13 merge and form a lateral edge of simple appearance”. Unfortunately, Kner only described the number of the coalescent plates, leaving out the last plates of the median series that are located at the base of the caudal peduncle (see Fig. 1 View FIGURE 1 ). Kner’s description included illustrations in lateral and dorsal views and a detail of the anteroventral region of the body ( Kner 1853:pl. 5).
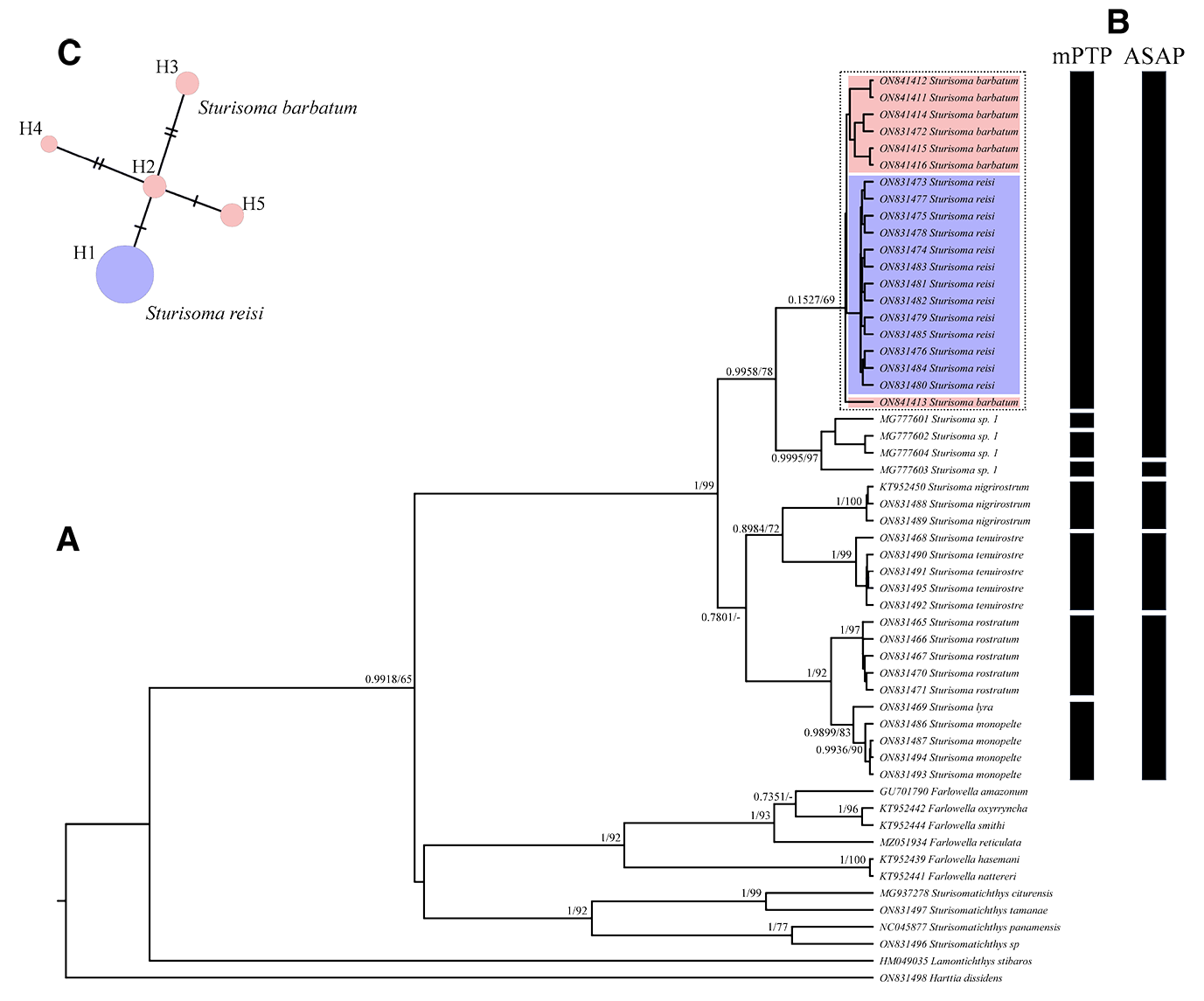
Later, Regan (1904) described S. robustum and compared it in detail with S. barbatum .We are not certain if Regan was able to examine the type-material of S. barbatum (NMW 46155), but the data suggests that he based many of his comparisons only on the artwork provided by Kner. Kner figures are artistically very thorough and allow us to clearly recognize the features that diagnose Sturisoma . However, when Kner’s illustrations are compared to the holotype of S. barbatum (NMW 46155), some inconsistencies may be noted: (i) the first two plates of the median lateral series were not delimited ( Figure 2B View FIGURE 2 ). Thus, if we count those first two plates in the illustration, the total number may be regarded as 34 (not including the supracaudal plate), however, a different number is seen on the holotype at hand (32) ( Fig. 1A–B View FIGURE 1 ). (ii) In the figure by Kner, the keels of the middle and lower series merge on plate 16 to continue into a single keel, while in the holotype this fusion is on plate 18 ( Fig. 1 A–B View FIGURE 1 ). (iii) In the figures, the dorsal fin originates before the ventral fins, whereas in the holotype the origin of the dorsal fin is just above the ventral fins ( Fig. 1 A–B View FIGURE 1 ). We assume that illustrative representations of S. barbatum ( Kner 1853:pl. 5), and not the first-hand analysis of the type specimen, conditioned the observations of Regan and subsequent authors, influencing the taxonomic history of S. barbatum . Regan (1904) affirmed that the distinction between S. barbatum and S. robustum in plate number where the keels of the middle and lower series join to continue in a single keel it is at the 16 for S. barbatum (15 + 17 + supracaudal = 33) and at 18 for S. robustum (17 + 15 + supracaudal = 33). However, the comparison of both holotypes results in the same number of plates: 17 + 15 + supracaudal = 33 for the midline. Kner (1853) indicated that the first ray of the dorsal fin in S. barbatum is rather thin and extends into a short thread, a feature not considered diagnostic by Regan (1904). The dorsal first ray of the type series of S. robustum is not more prominent than in S. barbatum . Our results indicate that these traits are variable and differ greatly according to ontogenetic stages, not being appropriate to differentiate these two species.
Another discussion based on the inaccuracies of Kner’s graphic representation includes that of Boeseman (1971) who questioned Bleeker’s (1862) description of the genus Oxyloricaria , who pointed out that the origin of the dorsal fin was just above the pelvic ones. In this line, Boeseman (1971: 5) mentioned that “it is only necessary to look at the illustrated figure” (with interpretation errors) of Kner to disprove Bleeker description. However, the review of the holotype specimen of S. barbatum indicates that Bleeker’s observation was right about this character. Otherwise, the graphic representation of S. robustum provided by Regan shows that his plate counting coincides completely with the type-material (see Fig. 2 C View FIGURE 2 ). We do not intend to discredit the drawings provided by Kner, nor Regan’s work, but to emphasize that the early graphic representations perpetuate to some degree the inconsistencies in the original diagnoses of these species.
However, in Londoño-Burbano & Reis (2021), using molecular and morphological data, the same authors showed that the sister group of Sturisoma is a clade composed of Sturisomatichthys + Farlowella + Aposturisoma . Our results, based only on one mitochondrial marker, support this last relationship between the genera of the tribe. Londoño-Burbano & Reis (2021) established that the genus Sturisoma includes only species with a cis-Andean distribution; and in their analyses, the distance between S. robustum and S. barbatum is surprisingly high. However, it should be taken into consideration that the S. barbatum specimen used in Londoño-Burbano & Reis (2021) analysis comes from the Madeira River (Amazon), a locality that is not included in the geographical distribution of the species and consequently, the specimen of S. barbatum analyzed by them may be misidentified.
New species of Sturisoma have been described in recent years, providing more information on S. barbatum . Ghazzi (2005) points out that this species has a moderate ventral expansion of the sphenotic and it is distinguished from most congeners by having 18 ventromedial plates without fused keels vs. 20–22 (usually 21) in other Sturisoma . By reviewing the type-material of S. barbatum and S. robustum , and the material from Argentina, we noticed the occurrence of 18 plates in the ventromedial series, and only two specimens (BMNH 1934.8.20.389–391 and CFA-IC-5103) had 19 and 20 plates without fused keels (see Fig. 5 View FIGURE 5 ) followed by 14 plates with fused keels (19+14+supracaudal=33). Regarding the sphenotic shape, we found in the holotype of S. barbatum and S. robustum that the sphenotic ventral expansion is absent in both taxa, which contradicts Ghazzi (2005; Londoño-Burbano, 2018) observation. Therefore, we estimate that the above-mentioned characters cannot be diagnostic to differentiate these two Sturisoma species.
In the key to differentiate Sturisoma species, Londoño-Burbano (2018) proposed the lengths of snout and of the distal filament that continues after the dorsal spine and the pectorals as characters to differentiate S. barbatum from S. robustum . We tested this proposal, but we found that these features are extremely variable because they are fragile and easily lost during the life of each specimen.
Londoño-Burbano & Britto (2022) recently described another species, S. reisi (from Madeira River, Bolivia, and Brazil), and suggested it is sister to S. barbatum , even the branch between both species was not well supported. Our molecular results did not distinguish these two species. The tree reconstruction, genetic distances, and delimitation methods suggested that specimens of S. barbatum and S. reisi belong to the same lineage. Despite a low genetic distance between S. barbatum and S. reisi (~0.2%, according to their data), Londoño-Burbano & Britto (2022) assumed the morphological evidence justified the distinction between both taxa. Despite the fact that our contribution aims to elucidate the status of La Plata Basin species, the fact that in our analysis S. reisi shows low genetic distance with S. barbatum , invites us to overview the morphological distinctiveness between both taxa. Londoño-Burbano & Britto (2022) supported that S. reisi can be distinguished by having a middorsal longitudinal, thin dark stripe on the caudal peduncle, extending from the second or third plate posterior to the dorsal-fin base to the origin of the caudal fin; central abdominal plates without spots ( vs. central abdominal plates with faded dark spots); anteriormost abdominal plates, just posterior to the lower lip, square, not enlarged, smaller than anterior abdominal plates ( vs. rectangular, enlarged, longer than anterior abdominal plates; presence of a small blotch on distal tips of the first two branched rays of anal fin ( vs. absence); and presence of poorly-developed odontodes at the sides of head on sexually dimorphic males ( vs. well-developed odontodes). As indicated in the descriptive section, and as noted in the figures, some of these features are also present in several S. barbatum specimens. However, some traits (i.e., coloration of caudal peduncle, abdominal plates, and anal fin, plus the dimorphic development in mature males on both species) may distinguish both forms as supported by Londoño-Burbano & Britto (2022).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Loricariinae |
|
Tribe |
Farlowellini |
|
Genus |
Sturisoma barbatum ( Kner, 1853 )
| Cardoso, Yamila, Bogan, Sergio, Agnolin, Federico & Petean, Flávia De Figueiredo 2023 |
Sturisoma robusta
| Ringuelet 1967 |
Oxyloricaria robusta
| Pozzi 1945 |
Oxyloricaria barbata
| Pozzi 1945 |
Loricaria barbata
| Kner 1853: 12 |