
Nogunius, Jałoszyński, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4822.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:86DC1241-FE11-49CC-9674-ACAB1DF92F62 |
|
DOI |
https://doi.org/10.5281/zenodo.4450827 |
|
persistent identifier |
https://treatment.plazi.org/id/525F87CC-FF90-FF98-9BED-FF75FBCFFB57 |
|
treatment provided by |
Plazi |
|
scientific name |
Nogunius |
| status |
gen. nov. |
Nogunius gen. n.
Type species: Nogunius sokani , here designated.
Diagnosis. Body elongate, with distinct constriction between head and pronotum and between pronotum and elytra; head short and round, with eyes situated anteriorly, adjacent to antennal fossae, and with long tempora; thick bristles present on head and prothorax; antennal fossae narrowly separated at middle; antenna with large, sharply delimited dimerous club; submentum with lateral sutures continuous with gular sutures; hypostomal ridges incomplete, running only slightly obliquely to lateral margins of cardines and reaching neither submentum nor posterior tentorial pits; pronotum subconical, broadest shortly in front of base and narrowing anterad, with rounded sides; pronotum with two pairs of antebasal pits; prosternum lacking prosternal process, with basisternal portion not demarcated; notosternal sutures distinct in front of procoxal cavities, but postcoxal process of hypomeron firmly fused with postcoxal process of coxosternum without a trace of suture, so that procoxal cavities are closed; hypomeral ridges absent; mesoventral intercoxal process carinate, broad; pleural structures of meso- and metaventrite partly fused laterad mesocoxal projections; metaventral intercoxal process subtrapezoidal, short, with median notch, narrowly separating metacoxae; each elytron with one small but distinct and asetose basal fovea; aedeagus with symmetrical median lobe and endophallus, and with free, slender parameres; spermatheca elongate.
Description. Body form ( Figs 45–51 View FIGURES 45–52 ) Euconnus -like, elongate, moderately slender, with distinct constrictions between head and pronotum and between pronotum and elytra.
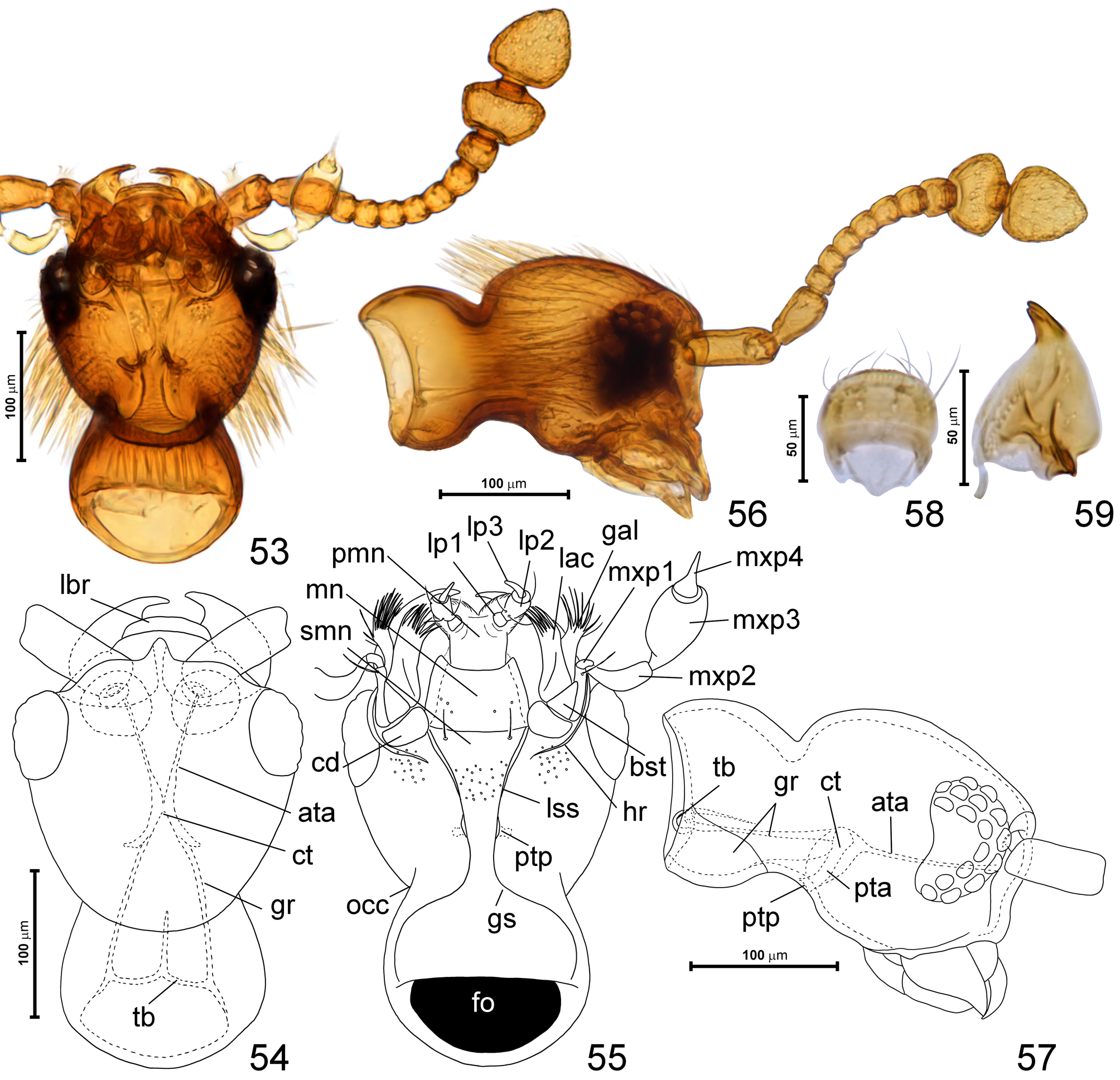
Head ( Figs 46, 48, 50, 52–57 View FIGURES 45–52 View FIGURES 53–59 ) round and strongly convex; eyes situated closer to mandibular bases than to occipital constriction; vertex not bulging posterodorsad, confluent with frons; frontoclypeal groove present, short; tempora and posterior margin of frons with thick, long bristles. Occipital constriction ( Fig. 55 View FIGURES 53–59 ; occ) about as broad as frons; ‘neck’ region slightly broadening posterad. Antennal fossae large and situated anterolaterally, narrowly separated at middle. Gular plate transverse, with distinct gular sutures marked on ‘neck’ region ( Fig. 55 View FIGURES 53–59 ; gs), anteriorly confluent with complete lateral sutures of submentum ( Fig. 55 View FIGURES 53–59 ; lss); posterior tentorial pits ( Fig. 55 View FIGURES 53–59 ; ptp) small, elongate, adjacent to gular sutures. Submentum ( Fig. 55 View FIGURES 53–59 ; smn) subtrapezoidal; mentum ( Fig. 55 View FIGURES 53–59 ; mn) subtrapezoidal with concave anterior margin; prementum ( Fig. 55 View FIGURES 53–59 ; pmn) largely membranous, flexible, lacking ligula, with moderately broadly separated bases of labial palps and small, inconspicuous lateral lobes of hypopharynx covered with microtrichia; labial palpomere I ( Fig. 55 View FIGURES 53–59 ; lp1) short, ring-like, palpomere II ( Fig. 55 View FIGURES 53–59 ; lp2) elongate, subcylindrical, palpomere III ( Fig. 55 View FIGURES 53–59 ; lp3) narrow, elongate, tapering distad and slightly curved. Maxilla ( Fig. 55 View FIGURES 53–59 ) with large cardo ( Fig. 55 View FIGURES 53–59 ; cd), subtriangular basistipes ( Fig. 55 View FIGURES 53–59 ; bst), broad mediostipes with an oblique groove demarcating elongate lacinia with a row of robust setae along mesal and distal margin, galea ( Fig. 55 View FIGURES 53–59 ; gal) elongate, with robust setae along distal and distomesal margin, palpifer ( Fig. 55 View FIGURES 53–59 ; ppf) elongate, maxillary palp with weakly elongate palpomere I ( Fig. 55 View FIGURES 53–59 ; mxp1), long, pipe-like palpomere II ( Fig. 55 View FIGURES 53–59 ; mxp2), broad and relatively short palpomere III ( Fig. 55 View FIGURES 53–59 ; mxp3), and subconical palpomere IV ( Fig. 55 View FIGURES 53–59 ; mxp4) with distal half nearly rod-like. Mandibles ( Figs 54, 59 View FIGURES 53–59 ) symmetrical, each subtriangular, with broad basal portion and narrow apical tooth, which is indistinctly bifurcate; setose prostheca present but short and indistinct. Labrum ( Fig. 58 View FIGURES 53–59 ) transverse, with rounded anterior margin, and with sparse, symmetrically distributed dorsal setae directed anterad.
Tentorium ( Figs 54, 57 View FIGURES 53–59 ) with small corporatentorium ( Figs 54, 57 View FIGURES 53–59 ; ct), bisinuate tentorial bridge ( Figs 54, 57 View FIGURES 53–59 ; tb) with anteromedian projection, long gular ridges ( Figs 54, 57 View FIGURES 53–59 ; gr; with membranous ventral portions and sclerotized dorsal portion, the latter visible in lateral and dorsal view as long ‘arms’), lacking dorsal tentorial arms, with short and stout posterior tentorial arms ( Fig. 57 View FIGURES 53–59 ; pta), and long, slender anterior tentorial arms ( Figs 54, 57 View FIGURES 53–59 ; ata).
Antenna ( Figs 53, 56 View FIGURES 53–59 ) slender, with scape and pedicel each strongly elongate, short and compactly assembled antennomeres III–VIII, slightly enlarged antennomere IX, and large, sharply delimited dimerous club, with antennomere XI slightly narrower than X.
Prothorax in dorsal view ( Figs 45–51 View FIGURES 45–52 ) subconical, broadest shortly in front of base but relatively weakly narrowing anterad. Anterior pronotal corners distinct, posterior corners broadly rounded. Pronotal base with two pairs of distinct pits. Sides of pronotum with thick bristles. Prosternum ( Figs 60–61 View FIGURES 60–63 ) short in relation to prothorax, with basisternal portion not demarcated and lacking prosternal process, laterally in front of procoxal cavities separated from hypomera by notosternal sutures ( Fig. 61 View FIGURES 60–63 ; nss), posteriorly lateral process of coxosternum firmly fused with postcoxal process of hypomeron, closing procoxal cavities; hypomeral ridges absent.
Mesonotum ( Fig. 62 View FIGURES 60–63 ) with mesoscutum (Fig, 62; sc2) and mesoscutellum ( Fig. 62 View FIGURES 60–63 ; scl2) demarcated by scutoscutellar suture ( Fig. 62 View FIGURES 60–63 ; sss); only tiny posterior area of scutellar shield visible between elytral bases in intact specimens. Hind wings fully developed, functional.
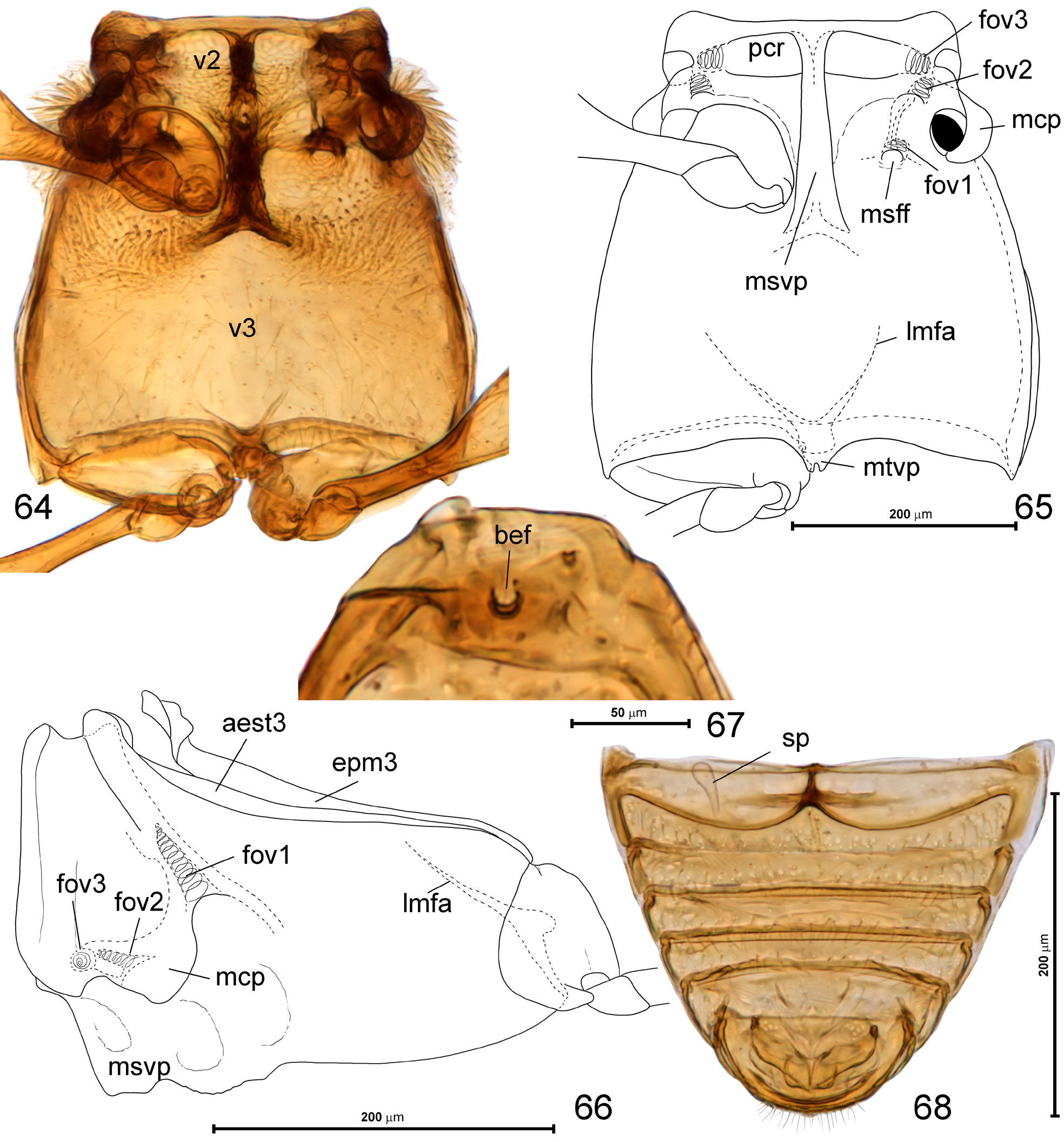
Mesoventrite ( Figs 64–66 View FIGURES 64–68 ) transverse, with short prepectus, transverse and setose anteromedian impressions functioning as procoxal rests ( Fig. 65 View FIGURES 64–68 ; pcr) separated at middle by long, carinate mesoventral intercoxal process ( Fig. 65 View FIGURES 64–68 ; msvp), which anteriorly nearly reaches anterior margin of mesoventrite. Two shallow foveae can be seen on sides of mesoventrite, each opening on dorsal surface of ventrite (i.e., not opening outside, but inside thorax): fovea 2 ( Fig. 65 View FIGURES 64–68 ; fov2) with opening at base of mesocoxal projection near tip of mesofurca, and fovea 3 ( Fig. 65 View FIGURES 64–68 ; fov3) opening on anteropleural region at the level of lateral corner of procoxal rest. Mesoventral pleural structures partly fused with metaventrite laterad mesocoxal projection ( Fig. 66 View FIGURES 64–68 ); the latter ( Fig. 65 View FIGURES 64–68 ; mcp) prominent, strongly projecting laterad, with mesocoxal cavities situated on ventromesal region and exposed in ventral view.
Metanotum ( Fig. 63 View FIGURES 60–63 ) with narrow alacristae ( Fig. 63 View FIGURES 60–63 ; alc), large metascutum ( Fig. 63 View FIGURES 60–63 ; sc3) with oblique internal apodemes ( Fig. 63 View FIGURES 60–63 ; ap), and a broad median membranous area ( Fig. 63 View FIGURES 60–63 ; mma); metascutellum not recognizable; metapostnotum ( Fig. 63 View FIGURES 60–63 ; pn3) short. Elytra ( Figs 45, 47, 49, 51 View FIGURES 45–52 , 67 View FIGURES 64–68 ) together oval, each with small humeral callus and round basal impression with a single asetose basal elytral fovea ( Fig. 67 View FIGURES 64–68 ; bef).
Metaventrite ( Figs 64–65 View FIGURES 64–68 ) subrectangular, anteriorly fused with mesoventrite, lacking anterior metaventral process, with metaventral intermetacoxal process ( Fig. 65 View FIGURES 64–68 ; mtvp) short, subtrapezoidal, with median notch, narrowly separating metacoxae. Mesofurcal foveae ( Fig. 65 View FIGURES 64–68 ; msff) open in median area of each mesocoxal rest, base of mesofurcal arm bears a deep projection with fovea 1 ( Figs 65–66 View FIGURES 64–68 ; fov1) opening internally and its walls forming additional projection (apophysa) inside pterothorax. Metanepisterna ( Fig. 66 View FIGURES 64–68 ; aest23) and metepimera ( Fig. 66 View FIGURES 64–68 ; epm3) fully demarcated and narrow.
Legs slender, lacking peculiar features.
Abdomen ( Fig. 68 View FIGURES 64–68 ) subtriangular, with six exposed sternites (sternites III–VIII), suture between VII and VIII indistinct.
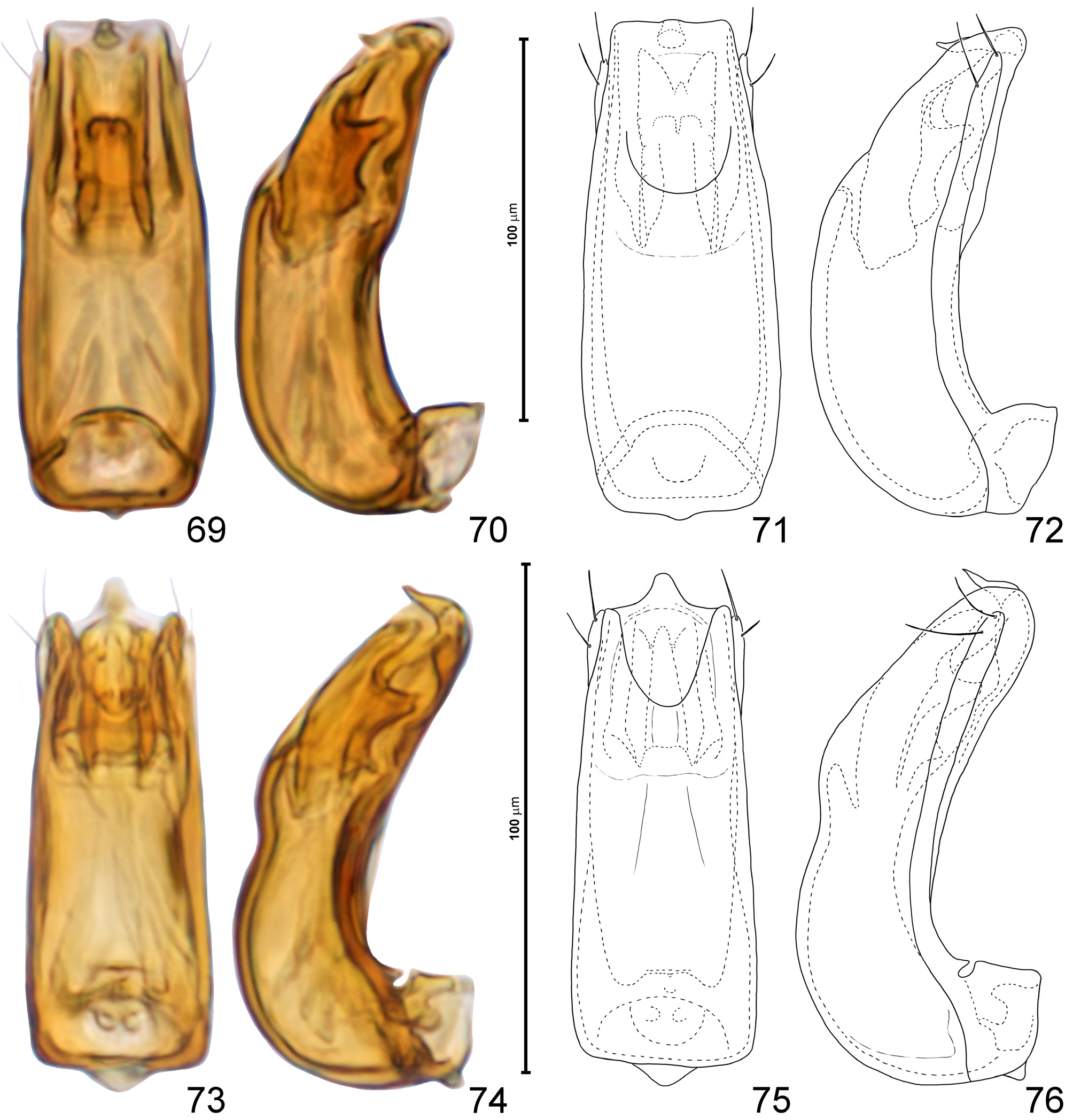
Aedeagus ( Figs 69–84 View FIGURES 69–76 View FIGURES 77–84 ) with symmetrical and nearly tubular median lobe with strongly curved basal region and basal foramen surrounded by large ‘collar’ formed of parameral bases; ostium situated ventrally in subapical region; endophallus symmetrical and composed of paired sclerites; parameres free (i.e., not fused with walls of median lobe), slender, each with apical and subapical setae.
Spermatheca ( Fig. 68 View FIGURES 64–68 ; sp) small, thin-walled and elongate.
Sexual dimorphism distinctly expressed only in size of eyes (smaller in females).
Etymology. This genus is named after Noguni Sôkan, an official who introduced the sweet potato to Okinawa from China in the beginning of the seventeenth century. The sweet potato then spread from Okinawa to Japan, and in the Ryukyus it became a staple food that saved many lives from famine. It is not a personal name; sôkan was a manager-level position in the Ryukyu Kingdome, and Noguni is a district of the Kadena town on Okinawa-jima, where he was born; the true name of Noguni Sôkan remains unknown. Gender masculine.
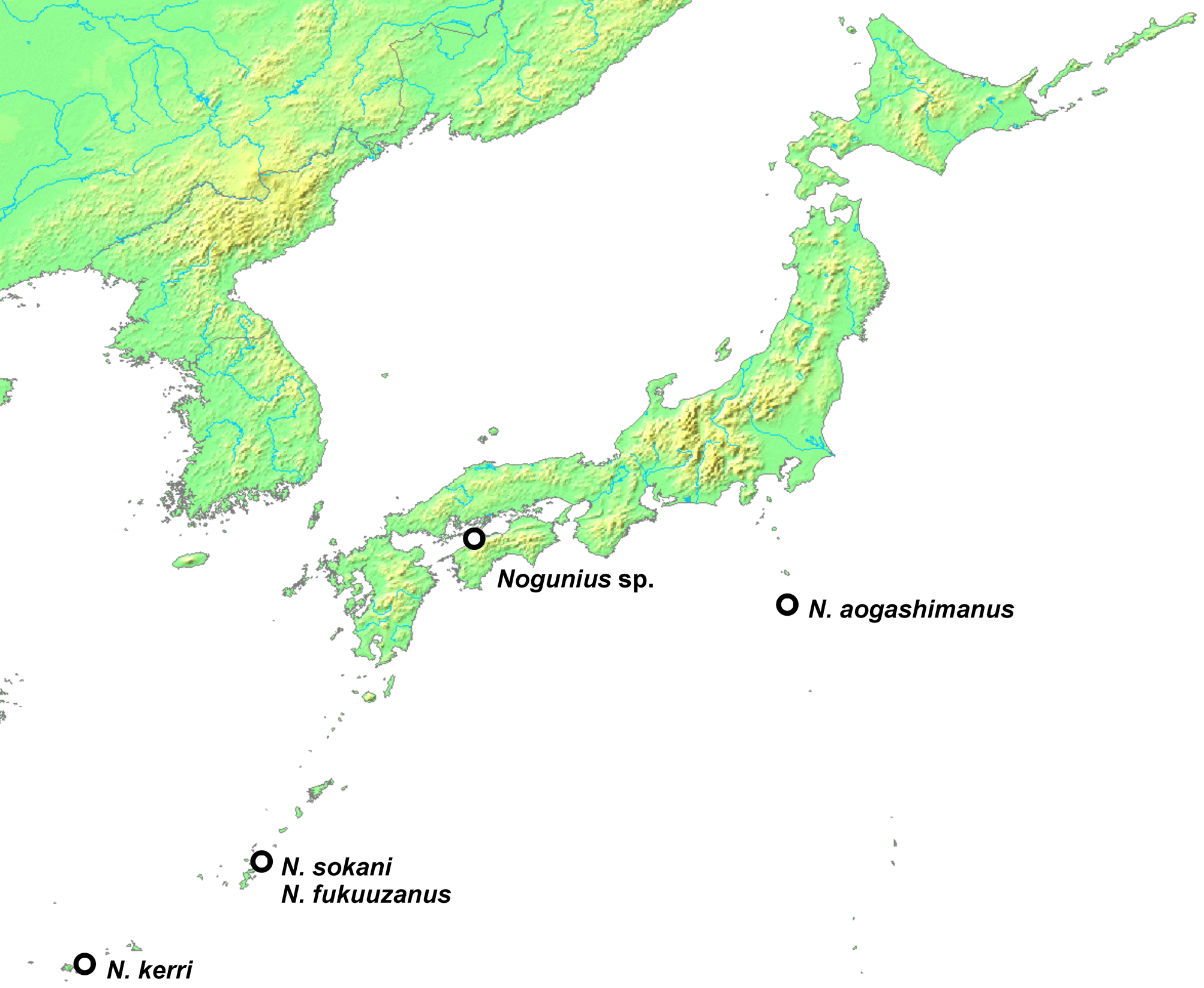
Composition and distribution. Nogunius is known to occur in the Ryukyus and Izu Islands; one female representing most likely an undescribed species was found on Shikoku ( Fig. 85 View FIGURE 85 ).
Remarks. Although adults of this genus resemble Euconnus and similar genera of Stenichnini , they have an unusual feature, so far within this tribe known only in Stenichnus -like genera, i.e., complete lateral sutures of the submentum. In Nogunius , the sutures are clearly identifiable as anterior (pre-tentorial) portions of gular sutures, with posterior tentorial pits situated on their lateral margins far in front of the transverse gular plate on the ‘neck’ region. The gular ridges extend along gular sutures inside the head capsule. Unambiguous homologization of the lateral sutures of the submentum with gular sutures was so far problematic, because the submental lateral sutures in Stenichnus -like genera meet the sutures of the posterior (post-tentorial) portion of the gular plate at some angle, and the posterior tentorial pits are hidden inside a deep transverse groove between the ‘neck’ region and the anterior, exposed part of the head capsule (e.g., Jałoszyński 2013). Consequently, each lateral suture could not be unambiguously interpreted as a single, continuous structure extending from the anteroventral margin of the foramen occipitale up to the posterolateral margin of the mentum. The continuity of the suture on the entire ventral surface of the head capsule in Nogunius seems undisputed, and this arrangement is similar to the gular sutures found in many other Staphylinidae (e.g., Solodovnikov & Newton 2005: fig. 2.).
Among all genera of Stenichnini , morphological structures of four are unknown to me: Napochomorphus Franz , Neuraphomorphus Reitter , Psepharobius King , and Taphroscydmus Casey. Each of them includes one species. Despite long efforts, I was not able to borrow the type material of the type species of the Brazilian Napochomorphus ; depositories of the type material of the type species of the Ethiopian Neuraphomorphus , the Australian Psepharobius and the North American Taphroscydmus remain unknown (none is present in Hungarian, Australian, and Russian museums that hold major parts of what remained from collections of Reitter, King and Motschulsky, respectively). The remaining genera can be divided into two groups based on the presence or absence of the lateral sutures of the submentum (the term “pre-tentorial gular sutures” will be used consistently in this and future studies). These groups may not be monophyletic, but such a preliminary division greatly facilitates identifications by excluding a large number of genera using a relatively easily observable character. The pre-tentorial gular sutures are known to occur in 23 genera, including Nogunius . Additionally, a re-assessment of cephalic structures in all specimens studied so far by scanning electron microscopy or as transparent mounts revealed that in Magellanoconnus s. str. structures interpreted previously as hypostomal ridges may in fact be the pre-tentorial gular sutures (discussion concerning the diagnosis and taxonomy of this genus will be included in a separate, phylogenetic paper; Jałoszyński in prep.). All genera with the pre-tentorial gular sutures can be divided into two groups:
1. The mesoventral intercoxal process absent: Austrostenichnus Franz , Neladius Casey , Palaeoscydmaenus Franz , and Stenichnaphes Franz.
2. The mesoventral intercoxal process or carina present: Alloraphes Franz , Brachycepsis Brendel , Catalinus Casey , Delius Casey , Leptoderoides Croissandeau , Madagaphes Jałoszyński , Magellanoconnus Franz , Mexiconnus Jałoszyński, Neuraphes Thomson , Nogunius Jałoszyński , Obesoconnus Jałoszyński , Parascydmus Casey , Rutaraphes Jałoszyński, Scydmaenilla King , Scydmoraphes Reitter , Siamites Franz , Stenichnodes Franz , Stenichnus Thomson , and Zeanichnus Jałoszyński.
The group 2 can be divided into two subgroups:
2A. Eyes clearly closer to occipital constriction than to mandibular bases: Alloraphes , Brachycepsis , Leptoderoides, Madagaphes , Neuraphes , Parascydmus , Rutaraphes , Scydmoraphes , Stenichnodes , Stenichnus , and Zeanichnus .
2B. Eyes either closer to mandibular bases than to the occipital constriction, or at middle, or so large that touching or nearly touching both mandibular bases and occipital constriction; or the placement of eye is ambiguous or variable within genus (e.g., because of the occurrence of macrophthalmous species with eyes close to middle and microphthalmous with eyes close to mandibular bases): Catalinus (eyes near middle), Delius (eyes near middle), Magellanoconnus s. str. (eyes clearly closer to mandibular bases), Mexiconnus (eyes slightly closer to mandibular bases), Nogunius (eyes clearly closer to mandibular bases), Obesoconnus (huge eyes occupying entire lateral surface of head), Scydmaenilla (eyes closer to occipital constriction or near middle), Siamites (eyes near middle, or slightly closer to mandibular bases, in microphthalmous specimens adjacent to antennal fossae and clearly closer to mandibular bases).
Within group 2B, Nogunius can be distinguished from all remaining genera by (1) the sharply delimited, dimerous antennal club; (2) antennal fossae narrowly separated at middle by distance narrower than width of scape; (3) anterior margin of the mentum distinctly concave; and (4) the pronotum broadest shortly in front of base. Additionally, Nogunius differs in several features from each of these genera:
- from Catalinus in thick bristles present on head and pronotum (lacking in Catalinus ); tempora strongly narrowing posterad and forming an even arc with posteriorly convex posterior margin of vertex (in Catalinus , tempora weakly narrowing posterad and delimited by distinct obtuse-angled corners from anteriorly convex posterior margin of vertex); posterior tentorial pits exposed and situated far in front of broad, diffuse transverse impression demarcating ventrally ‘neck’ region (in Catalinus posterior tentorial pits hidden in deep, narrow, sharply marked transverse groove demarcating ventrally ‘neck’ region); hypostomal ridges incomplete, short, developed only shortly behind cardines (in Catalinus hypostomal ridges complete, long, posteromesally confluent with transverse groove in which posterior tentorial pits are situated); pronotum with four antebasal pits (pits lacking in Catalinus ); hypomeral ridges lacking (in Catalinus incomplete but present); posteromedian margin of prothoracic coxosternum with shallow emargination (in Catalinus with subtriangular projection); basal elytral fovea situated near middle of elytral articulating lobe, distant from sutural margin (in Catalinus close to sutural margin); mesoventral intercoxal process broad and behind mesocoxae indistinguishably fused with metaventrite (in Catalinus process narrow and reaching only to middle of mesocoxae, where it forms a distinct posterior tip separated from metaventrite); and in symmetrical endophallic structures (in Catalinus asymmetrical);
- from Delius in tempora strongly narrowing posterad and forming an even arc with posteriorly convex posterior margin of vertex (in Delius , tempora weakly narrowing posterad and delimited by distinct obtuse-angled corners from anteriorly convex posterior margin of vertex); hypostomal ridges incomplete, short, developed only shortly behind cardines (in Delius hypostomal ridges complete, long, posteromesally reaching posterior tentorial pits); pronotum with inner antebasal pits not connected by transverse groove (groove present in Delius ); hypomeral ridges lacking (in Delius present); basal elytral fovea situated near middle of elytral articulating lobe, distant from sutural margin (in Delius basal elytral fovea close to sutural margin); mesoventral intercoxal process with straight lateral margins gradually diverging from anterior to posterior region (in Delius process distinctly constricted between mesocoxae); and metacoxae laterally reaching sides of metaventrite (in Delius lateral corners of metacoxae are broadly separated from lateral margins of metaventrite);
- from Magellanoconnus s. str. in pronotum lacking sublateral carinae (present in Magellanoconnus ); hypomeral ridges lacking (in Magellanoconnus present); procoxal cavities closed (open in Magellanoconnus ); posteromedian margin of prothoracic coxosternum with shallow emargination (in Magellanoconnus with subtriangular projection); one basal fovea on each elytron (two in Magellanoconnus ); mesoventral intercoxal process long, behind mesocoxae fused with metaventrite (in Magellanoconnus process reaching only to middle of mesocoxae, where it forms a distinct posterior tip separated from metaventrite); and anterior metaventral process lacking (in Magellanoconnus present);
- from Mexiconnus in head with thick bristles (lacking in Mexiconnus ); pronotum lacking transverse antebasal groove (groove present in Mexiconnus ); hypomeral ridges lacking (in Mexiconnus present); each elytron with one distinct basal fovea (in Mexiconnus two barely discernible vestiges of foveae); protarsi in males unmodified (strongly broadened and with long and dense ventral setal ‘cushions’ in Mexiconnus ); and aedeagus with symmetrical endophallus and slender, free parameres bearing apical setae (in Mexiconnus endophallus asymmetrical and parameres in basal halves broader than median lobe, with asetose apices);
- from Obesoconnus in distinct tempora and distinct occipital constriction (both lacking in Obesoconnus ); head and pronotum with thick bristles (absent in Obesoconnus ); posterior tentorial pits exposed and situated far in front of broad, diffuse transverse impression demarcating ventrally ‘neck’ region (in Obesoconnus posterior tentorial pits hidden in a deep, narrow, sharply marked transverse groove demarcating ventrally ‘neck’ region); pronotum lacking transverse antebasal groove (groove present in Obesoconnus ); maxillary palpomere III strongly elongate and with distal half nearly rod-like (stout, evenly subtriangular in Obesoconnus ); procoxal cavities closed (open in Obesoconnus); hypomeral ridges absent (present in Obesoconnus ); posteromedian emargination of prothoracic coxosternum shallow, barely marked (in Obesoconnus conspicuously deep, reaching nearly half length of prosternum); mesoscutellar shield tiny and barely discernible between elytral bases (in Obesoconnus conspicuously large and well-visible); and aedeagus lacking basal ‘pumping apparatus’ (in Obesoconnus present, composed of basal diaphragm with sclerotized median area to which internal longitudinal apodeme with longitudinal muscles is attached);
- from Scydmaenilla in head with thick bristles (absent in Scydmaenilla ); posterior tentorial pits exposed and situated far in front of broad, diffuse transverse impression demarcating ventrally ‘neck’ region (in Scydmaenilla posterior tentorial pits are hidden in a deep, narrow, sharply marked transverse groove demarcating ventrally ‘neck’ region); pronotum with two pairs of antebasal pits, lacking transverse groove (in Scydmaenilla one pair of pits and transverse groove are present); postcoxal coxosternal and hypomeral processes firmly fused (in Scydmaenilla posterior hypomeral process overlaps coxosternal process, but margins of each are well discernible); mesoventral intercoxal process long, behind mesocoxae fused with metaventrite (in Scydmaenilla process reaching only to middle of mesocoxae, where it forms a distinct posterior tip separated from metaventrite); and anterior metaventral process lacking (in Scydmaenilla present);
- from Siamites in lacking paired, dense tufts of bristles on vertex, prosternum, meso- and metaventrite and posterior margins of abdominal sternites (in Siamites conspicuous tufts or rosettes of bristles present on each of these regions); pre-tentorial gular sutures separated at middle (in Siamites connected between posterior tentorial pits); incomplete, short hypostomal ridges (in Siamites complete, reaching level of posterior tentorial pits); pronotum lacking transverse antebasal groove (in Siamites present); procoxal cavities closed (open in Siamites ); hypomeral ridges lacking (present in Siamites ); and mesoventral intercoxal process behind mesocoxae fused with metaventrite (in Siamites process forms a distinct posterior tip separated from metaventrite).
As two genera are here reported to occur in Japan for the first time, the identification key to Japanese genera of Stenichnini ( Cyrtoscydmini then) given by Jałoszyński (2015) must be updated.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Scydmaeninae |