
Eucalyptus odorata, Behr, Behr
|
publication ID |
https://doi.org/ 10.1071/SB21029 |
|
DOI |
https://doi.org/10.5281/zenodo.10974611 |
|
persistent identifier |
https://treatment.plazi.org/id/5261160A-FF89-FFD4-BB67-43BFFBD3B7A2 |
|
treatment provided by |
Felipe |
|
scientific name |
Eucalyptus odorata |
| status |
|
Genetic variation within the core E. odorata View in CoL complex as a cline
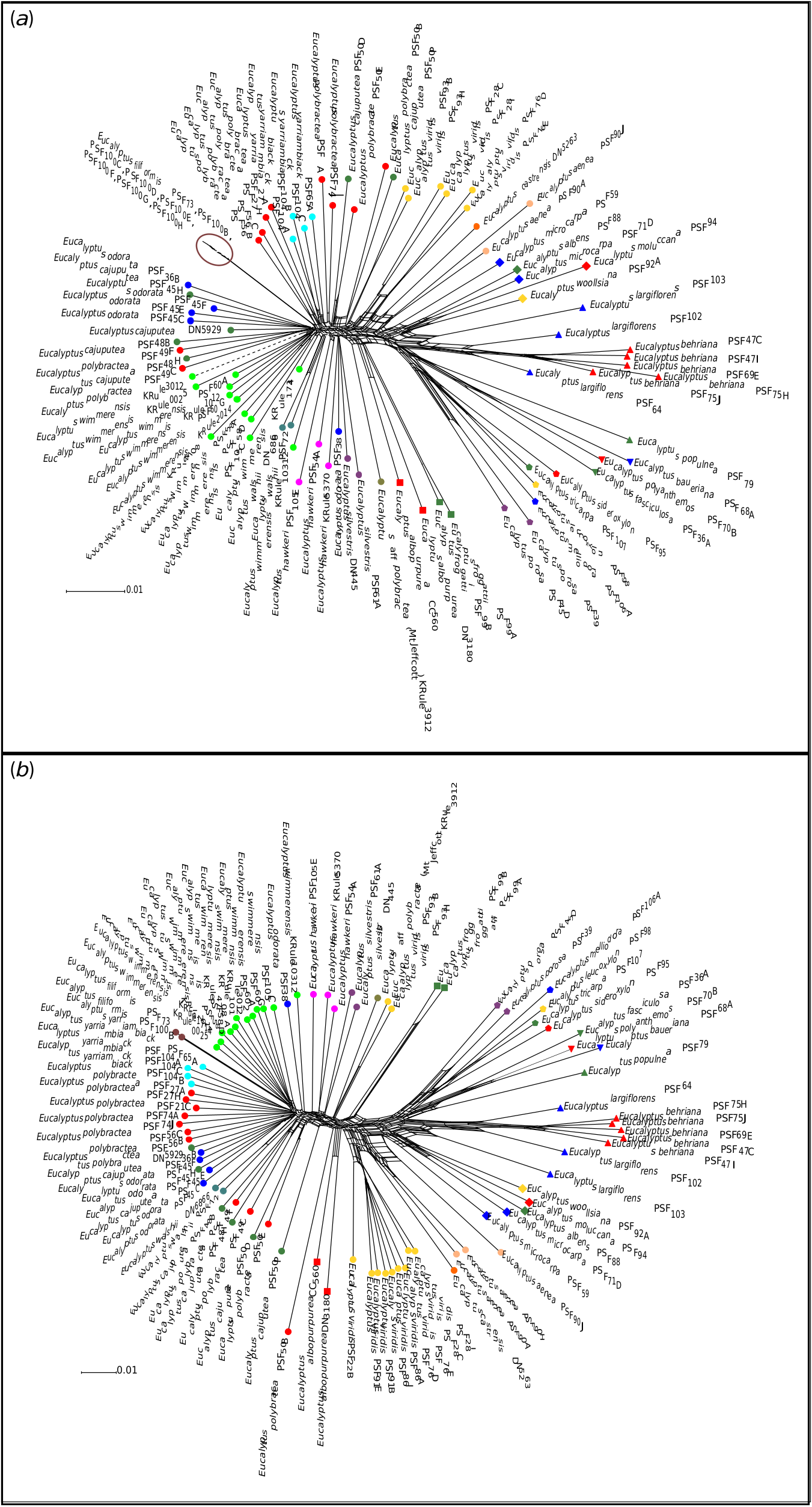
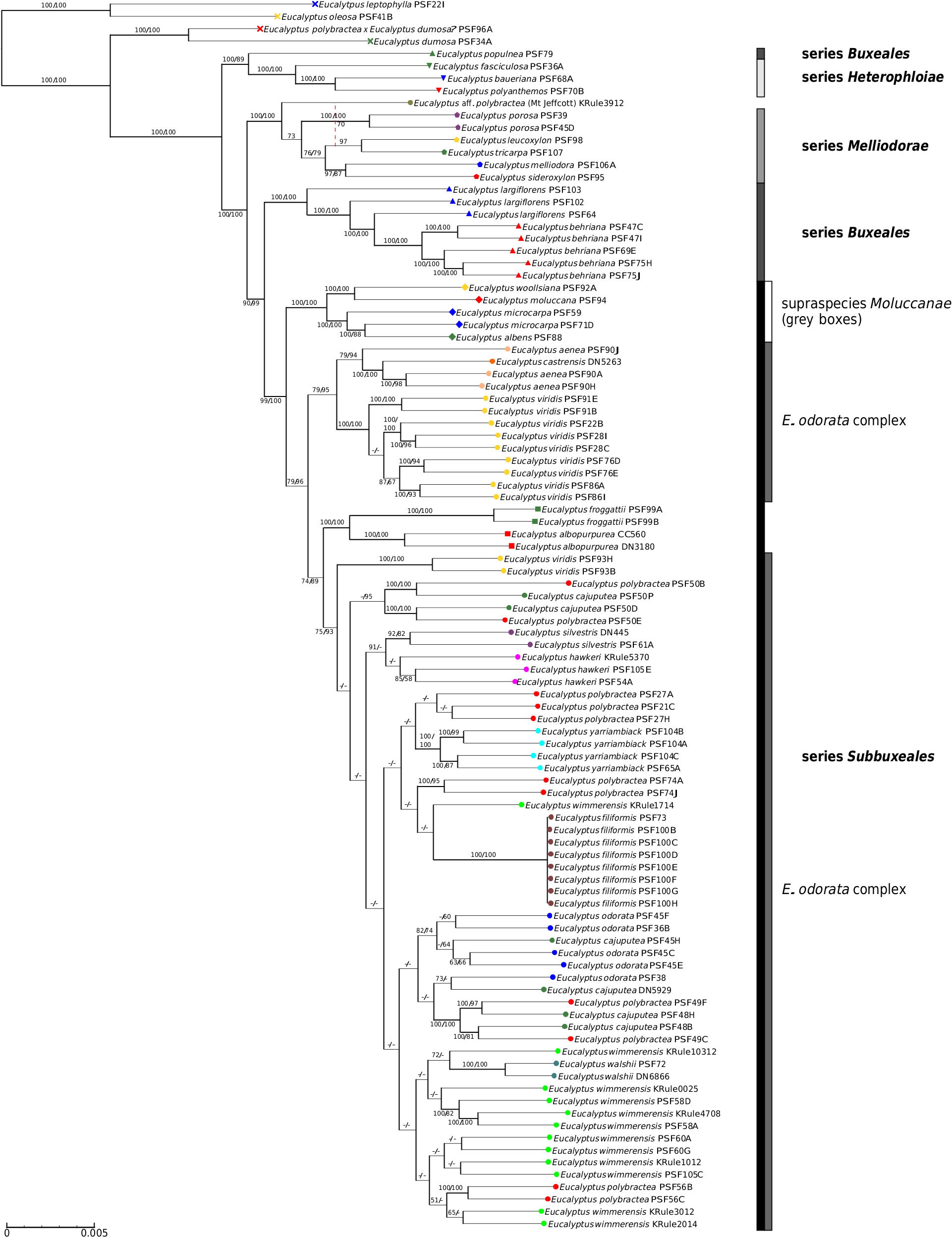
Our findings of extensive introgression among members of the E. odorata complex and co-occurring box species is not unexpected given that previous studies on E. section Adnataria have shown extensive hybridisation leading to morphological taxa not forming genetic clades ( Flores-Rentería et al. 2017). However, in regard to the taxonomy of the E. odorata complex, the nature of the core of the clade as a discontinuous cline of morphological and genetic variation running from the Flinders Ranges, south and then east through south-eastern SA, east through the Wimmera and then the Goldfields of Victoria and then north to West Wyalong in NSW also plays a significant role in disagreements among authorities over where taxonomic boundaries should be drawn. This clinal genetic variation has been taxonomically divided by different authors at different points on the basis of different factors they consider most important for classification and, in this paper, we have, a priori, broken it into the seven largely geographically distinct species ( E. odorata , E. cajuputea , E. wimmerensis , E. polybractea , E. filiformis , E. walshii and E. yarriambiack ).
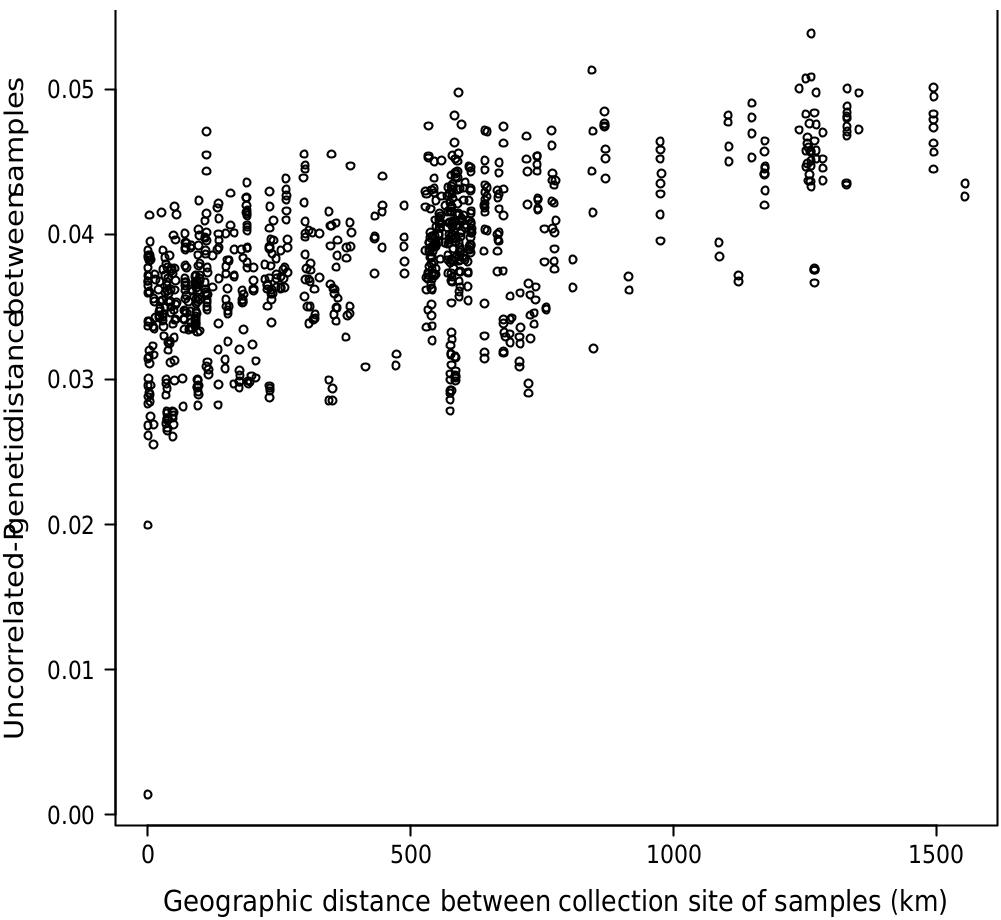
We see evidence for this cline in the most distal populations (Flinders Range and West Wyalong) being the most distinct, and in relationships between neighbouring populations fluctuate between analyses. This includes, for instance, E. filiformis clustering with E. wimmerensis in one network ( Fig. 2 b View Fig ) but being more closely related to E. polybractea from the Victorian goldfields in other analyses ( Fig. 2 a View Fig , 5), and the swapping of relationships between the three main groups (samples from west of the Murray River Basin, E. wimmerensis and allied samples, and eastern E. polybractea and allied samples) in the core group between analyses, even if the alternate relationships are not supported ( Fig. 4 View Fig ). This may be indicative of recent rapid diversification of a widespread ancestral population that has undergone vicariance at multiple locations at approximately congruent times. Clines along which diversification has occurred, often resulting in morphologically distinct species with a hybrid zone between them, have previously been observed in other eucalypt groups, including E. populnea and E. brownii Maiden & Cambage ( Holman et al. 2003), E. melanophloia F.Muell. and E. whitei Maiden & Blakely ( Holman et al. 2011), and the green ashes ( E. sect. Eucalyptus ; Rutherford et al. 2018). The strong support for IBD ( Fig. 6 View Fig , r: 0.597) in our data is congruent with the existence of a genetic cline.
| E |
Royal Botanic Garden Edinburgh |
| SA |
Museum national d'Histoire Naturelle, Laboratiore de Paleontologie |
| NSW |
Royal Botanic Gardens, National Herbarium of New South Wales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.