
DUNKLEOSTEIDAE STENSIÖ, 1963
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00578.x |
|
persistent identifier |
https://treatment.plazi.org/id/5415C76B-FF87-FFB1-9854-D1B0FC2C5093 |
|
treatment provided by |
Valdenar |
|
scientific name |
DUNKLEOSTEIDAE STENSIÖ, 1963 |
| status |
|
FAMILY DUNKLEOSTEIDAE STENSIÖ, 1963
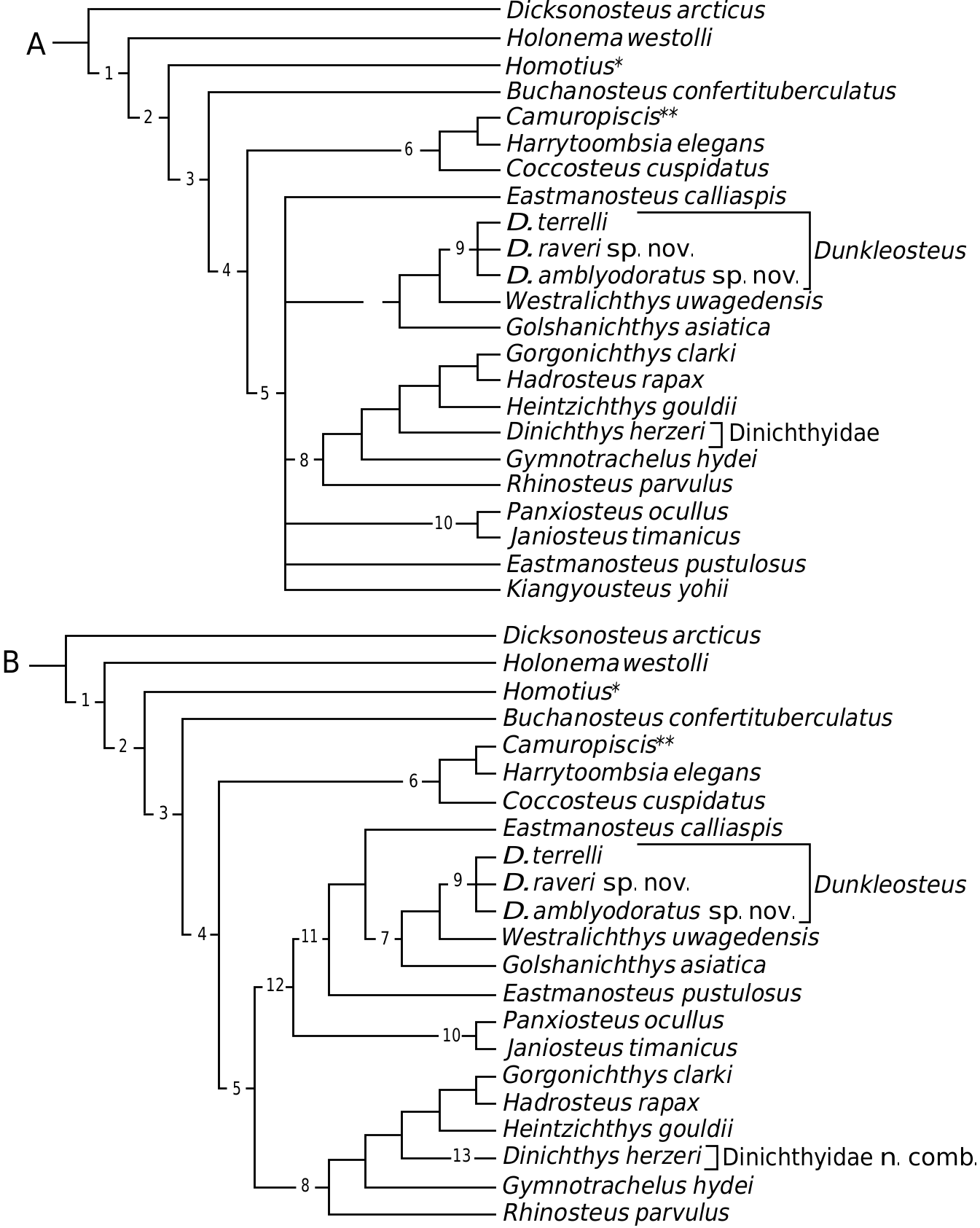
Stensiö (1963) erected the family Dunkleosteidae based on the members of Dunkle & Bungart’s (1946) terrelli group ( Du. terrelli , Du. intermedius , and Du. curtis , which were later synonymized with Du. prentis - clarki as Du. terrelli, Hlavin, 1976 ; Denison, 1978). Lelièvre et al. (1981) and Long (1987) added Gol. asiatica and W. uwagedensis , respectively, to the ‘dinichthyid’ arthrodires. Assuming that He. gouldii and Gor. clarki (which were included in these latter two studies) are more closely related to the other aspinothoracid arthrodires, the cladograms of Lelièvre et al. (1981, fig. 6) and Long (1987, fig. 11) suggest that the remaining ‘dinichthyids’ constitute a paraphyletic sequence.
The results from the present study do not support a paraphyletic status among these taxa ( Fig. 9 View Figure 9 ; see also Carr, 1991). Heintzichthys gouldii and Gor. clarki are members of the Aspinothoracidi, whereas Dunkleosteus , Ea. calliaspis , Ea. pustulosus , Gol. asiatica , and W. uwagedensis are members of the Dunkleosteidae . Dunkleosteus is diagnosed unambiguously by the development of a concave posterior margin on the nuchal plate (character 11, state 2). Several characters potentially unite Dunkleosteus species , although the missing data within individual Dunkleosteus species or in W. uwagedensis introduce some ambiguity: (1) the reduction or loss of the preorbital and paranuchal embayments of the central plate, and the development of a postorbital plate embayment of the central plate (character 13, state 0; character 14, state 0; and character 20, state 1; respectively); (2) the reduction or loss of the dermal postnuchal process on the paranuchal plate (character 30, state 0); and (3) the presence of anterior superognathal plate contact faces on the parasphenoid prehypophysial region (character 86, state 1). The potential data that might refute part of this diagnosis are the possible presence of parasphenoid articular facets ( Boylan, 1973) in Ea. pustulosus , Dinichthys cf. pustulosus (assumed to be a coccosteomorph by Stensiö, 1963: 386, footnote 1), and K. yohii ( Denison, 1978) . Boylan (1973: 396) interpreted the parasphenoid of Di. cf. pustulosus ( Kulczycki, 1956) to be of the Du. terrelli type, with ‘the concavities... much more subdued than those of Du. terrelli ’. We interpret from Kulczycki’s original figures ( Kulczycki, 1956: plate 1, figs G, H; plate 2, fig. 3) that the articular facets are absent. The prehypophysial region in Di. cf. pustulosus lacks any thickening associated with any contact face or articular facets. Furthermore, the parasphenoid in Di. cf. pustulosus is similar to that of Ea. calliaspis ( Dennis-Bryan, 1987, fig. 17); although, the prehypophysial region is wider in the former. The shape of the posterior process of the anterior superognathal in Ea. calliaspis supports the absence of a gnathal– parasphenoid contact in Eastmanosteus . Denison (1978) considered the parasphenoid of K. yohii to resemble that of Du. terrelli . Dennis-Bryan (1995) suggested that the orientation of the parasphenoid in K. yohii might be reversed (i.e. the assumed prehypophysial region should be considered as the posthypophysial region). This reversal was based on the position of the buccohypophysial depression. Dennis- Bryan noted that Denison’s (1978) interpretation places the buccohypophysial depression in an unlikely and unique anterior position. An alternative possibility is that the anterior region is incomplete (suggested by the uneven margin illustrated by Liu, 1955: plate II). In the current analysis, this feature is treated as an unknown.
Within Dunkleosteus , we considered three North American species ( Du. raveri sp. nov., Du. amblyodoratus sp. nov., and Du. terrelli ). Dunkleosteus raveri sp. nov. is nearly indistinguishable from Du. terrelli , with the former taxon characterized by a slight anterior shift of the anterior triple point (PtO/PrO/C plate junction) to a region over the orbit, suggesting either a shift in plate proportions or a relative enlargement of the orbit. The variability of this character within the Dunkleosteidae is not known, and may suggest that this feature provides little or no phylogenetic information. If the latter is true, Dunkleosteus raveri sp. nov. represents a metaspecies (a species that lacks any apomorphies, and thus cannot be ruled out as a possible ancestor; Archibald, 1994). Dunkleosteus raveri sp. nov. retains a number of primitive features, the most important being the retention of a dermal ornamentation (as noted above, the presence or absence of ornamentation in the Dunkleosteus species from the Ohio Shale Formation has a distinct stratigraphic distribution). Dunkleosteus amblyodoratus sp. nov., although fragmentary, is united with other Dunkleosteus species because of the presence of a contact between the parasphenoid and anterior superognathals. Dunkleosteus amblyodoratus sp. nov. lacks any unambiguous diagnostic features, thereby representing either a metaspecies or a potential ancestor of Du. terrelli and Du. raveri sp. nov. The anterior articulation of the parasphenoid in Dunkleosteus amblyodoratus sp. nov. may represent a transition to the more derived condition of transverse articular facets.
Eastmanasteus pustulosus is the sister taxon for ((( Dunkleosteus , W. uwagedensis ) Gol. asiatica ) Ea. calliaspis ). Thus, the genus Eastmanosteus represents a paraphyletic group ( Fig. 9B View Figure 9 ). It is noteworthy that the type specimen for Ea. pustulosus is considered as a nomen dubium by Carr (1991: 382). A revision of the Eastmanosteus species is hence required.
Another taxon that may be related to dunkleosteids is Pa. ocullus . The form of the prehypophysial region in Pa. ocullus ( Wang, 1979, fig. 2) is similar to Di. cf. pustulosus . In addition, the anterior superognathal posterior process lacks the enlargement seen in Du. terrelli , and is similar to that of Ea. calliaspis . In the specimen described by Wang, the parasphenoid appears to be anteriorly displaced, and hence does not represent a natural articulation: the prehypophysial region is in contact with the base of the anterior superognathal cusps; the anterior superognathal dorsal processes extend to the level of the buccohypophysial foramen (this position would have placed the hypophysis in the ethmoidal region of the chondrocranium, nearly between the two anterior superognathals).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |