
Lepidocyrtus florae Winkler & Mateos, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4429.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:AA87BBFC-DF19-4A05-A227-133C0A92049A |
|
DOI |
https://doi.org/10.5281/zenodo.5950499 |
|
persistent identifier |
https://treatment.plazi.org/id/5E0E41AD-83D5-4F75-87A4-F4BC759A5094 |
|
taxon LSID |
lsid:zoobank.org:act:5E0E41AD-83D5-4F75-87A4-F4BC759A5094 |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidocyrtus florae Winkler & Mateos |
| status |
sp. nov. |
Lepidocyrtus florae Winkler & Mateos View in CoL sp. nov.
Figs 11‒33 View FIGURE 11 View FIGURES 12–17 View FIGURES 18–19 View FIGURES 20–22 View FIGURES 23–24 View FIGURES 25–27 View FIGURES 28–33 , Tab 1
Type material. Holotype: female on slide (slide code: HNHM-collpr-807), Hanság , Csíkos Éger, com. Győr- Moson-Sopron ( Hungary), 124 m above sea level, N 47°40'47"; E 17°07'53", from moss on dead lying alder trees, hand collecting, 11.iv.2016, leg. P. Kelemen & D GoogleMaps . Winkler. Paratypes: 2 females and 1 male on slide (slide code: HNHM-collpr-808 to HNHM-collpr-810), and 3 specimens without visible genital plate on slides (code: LEP- FLO-001 to LEP-FLO-003); same data as holotype. The holotype and three paratypes (HNHM-collpr series) are deposited at the HNHM, Budapest. Three paratypes (LEP-FLO series) are saved in the collection of the author (D. Winkler) at the US, Sopron. GoogleMaps
Diagnosis. Small (1.1 mm of maximum length of body), specimens in alcohol evenly dark blue coloured ( Fig. 11 View FIGURE 11 ). Th.II not projecting over head. Antennae and legs (except for cx. II‒III and occasionally femur III) without scales. Apical bulb on ant.IV absent. Labial chaetae (M1M2REL1L2) in “p row” well developed and ciliated, R slightly shortened. Dorsal cephalic and body macrochaetae formula as A0A2a A2A3S3Pa5/00/0101+2.
Etymology. The species is named after D. Winkler’s niece Flora De Bonis.
Description. Holotype body length 1.05 mm (without head nor furca), paratypes 0.8‒1.1 mm. Th.II not projecting over the head ( Fig. 11a View FIGURE 11 ). Trunk and coxae uniformly dark blue ( Fig. 11a View FIGURE 11 ). Head apically blue, posteriorly white with blue marbling ( Fig. 11b View FIGURE 11 ). Ant. III‒IV blue, Ant. I‒II white on basal part but with increasing intensity of blue to distal part of segment. Blue shades also on manubrium and ventral tube. Scales densely covering head and trunk, coxae II‒III and femur III occasionally, and anterior side of manubrium and dens. Antennae and ventral tube without scales.
Antennae rather short, antennal length to head diagonal length ratio about 1.5 (head diagonal measured from cervical edge to apex of the mouth part). Relation of antennal joints I‒IV as 1:1.6‒1.8:1.5‒1.7:2.7‒3.1. Antennal base with two pseudopores on inner side and ventrolaterally with antennobasal lenticular organ (sensu Hüther 1986). Ant. III sense organ composed of two bent sensory rods partially behind a cuticular fold ( Fig. 12 View FIGURES 12–17 ). Ant. IV without apical bulb.
Arrangement of chaetae on the labrum 4/554, prelabral chaetae ciliated, all labral chaetae smooth ( Fig. 13 View FIGURES 12–17 ). Labrum intrusion inverted V-shaped, labral edge with four distinguishably separated, but not markedly protuberant rounded labral papillae, outer ones bigger in size ( Fig. 13 View FIGURES 12–17 ). Outer maxillary palp with two smooth chaetae ( Fig. 13 View FIGURES 12–17 ) and three smooth sublobal chaetae. Clypeus with twelve ciliated chaetae (3 in row pf, 5 in row f, and 2‒ 2 in rows L1‒L2, respectively) ( Fig. 14 View FIGURES 12–17 ). Labial palp with lateral process on papilla E slightly curved ( Fig. 15 View FIGURES 12–17 ), barely reaching top of papilla.
Labium chaetotaxy formed by 5 smooth chaetae (a1‒a5) in anterior row; basal row with ciliated chaetae M1M2REL1L2 ( Fig. 16 View FIGURES 12–17 ) with R somewhat smaller than other chaetae (ratio of R/M ~0.7). Ventral cephalic grove with 4+4 ciliated chaetae ( Fig. 16 View FIGURES 12–17 ).
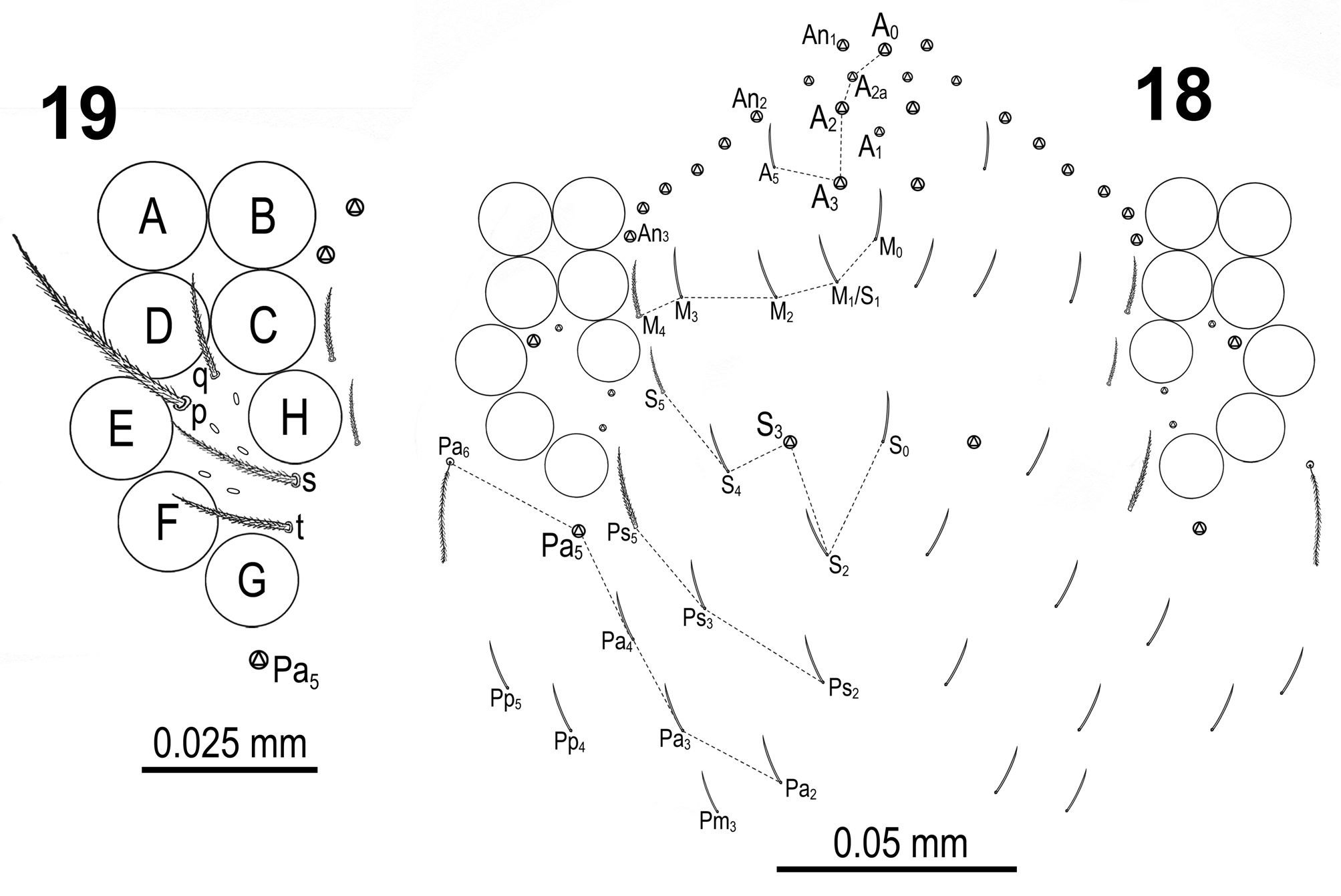
Dorsal cephalic macrochaetae chaetotaxy A0A2A3S3Pa5 ( Figs 17‒18 View FIGURES 12–17 View FIGURES 18–19 ), but also with pair of smaller supplementary macrochaetae A2a between A2 and A3; maximum number of macrochaetae An on head 8+8 ( Fig. 18 View FIGURES 18–19 ).
Eye patches dark blue, eyes well visible. Diameters of eyes A‒F about the same. Eyes G and H slightly smaller (A:G; A:H ≈ 1.1). Interocular chaetotaxy ( Fig. 19 View FIGURES 18–19 ) with s, t, p, q chaetae and 3‒5 intraocular scales.
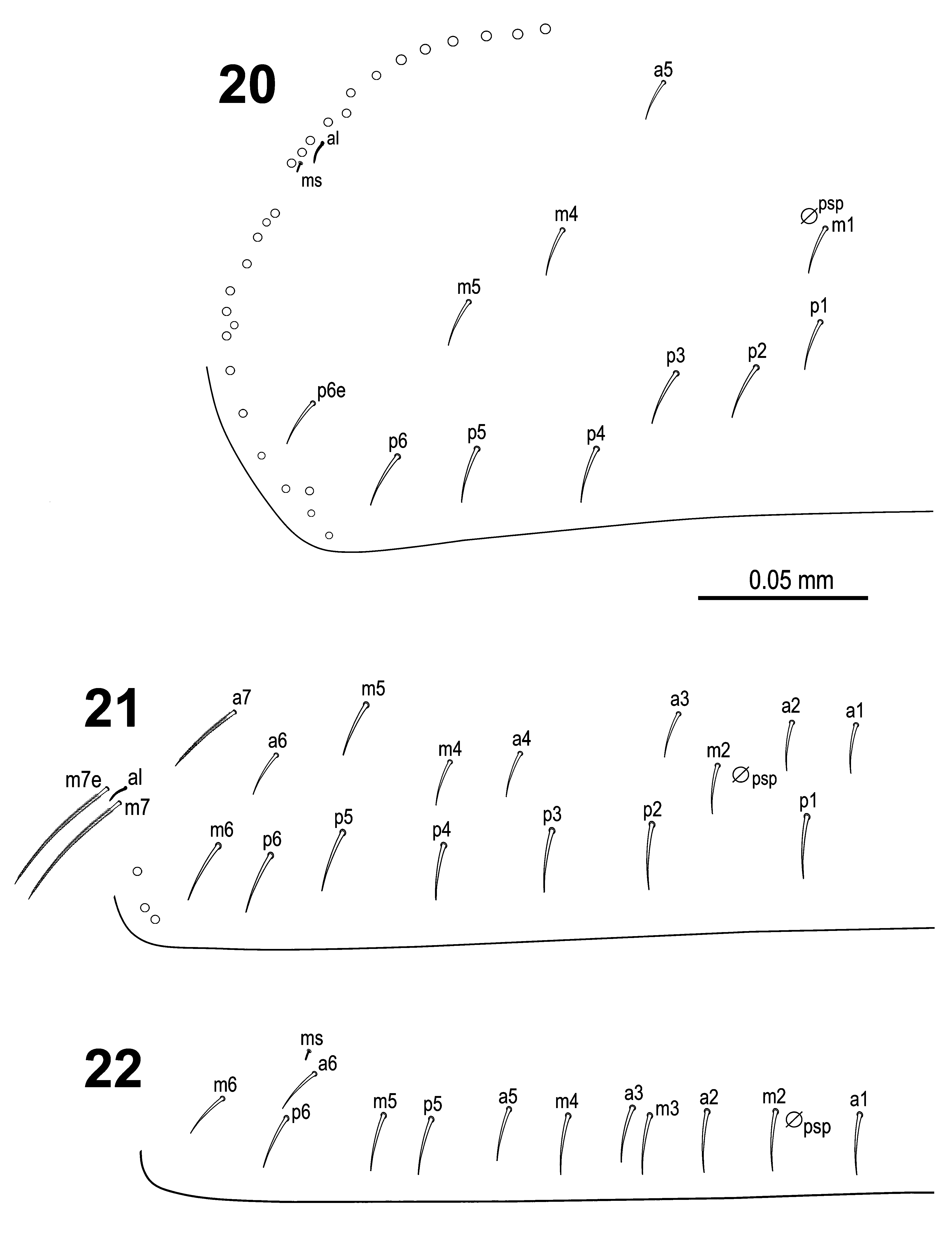
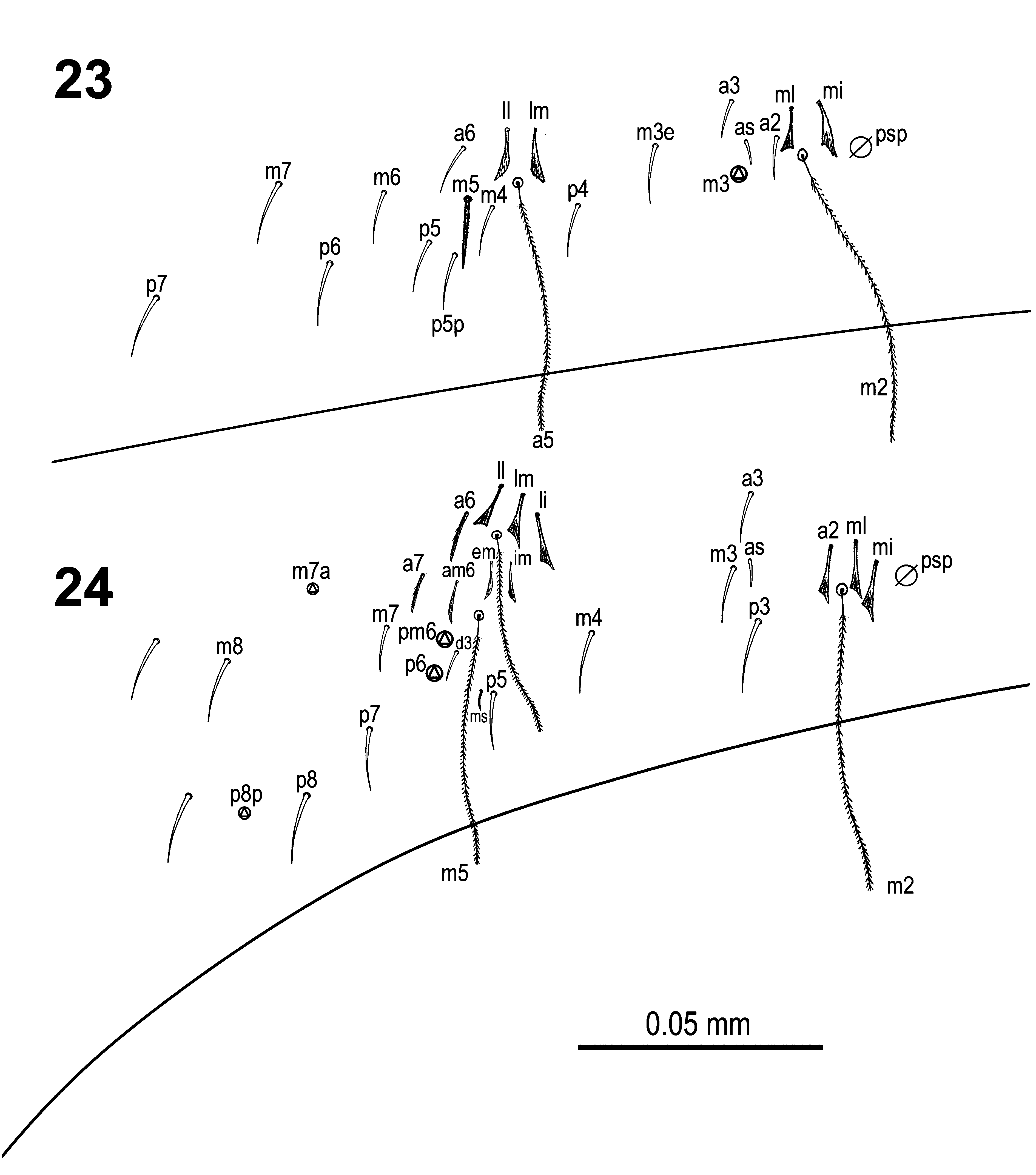
Body macrochaetae 00/0101+2 ( Fig. 17 View FIGURES 12–17 ). Dorsal chaetotaxy of th.II‒III and abd.I as in Figs 20‒22 View FIGURES 20–22 . Th.II with 2 anterolateral S-chaetae (al and ms) and without macrochaetae in posterolateral position. Th.III with a sensillum (al) guarded by ciliated chaetae m7 and m7e. Abd.I with a lateral S-microchaeta (ms) external to a6. Chaetotaxy of abd.II‒III as in Figs 23‒24 View FIGURES 23–24 . Abd.II chaetotaxy between the two dorso-medial trichobothria characterized by mesochaetae a2, a3, m3e, p4, sens as and macrochaeta m3; chaeta a2p absent; chaeta m5 very thin and short, spine-like slightly ciliated macrochaeta (ratio of macrochaeta m3 to m5: 4.6‒5.4). On abd.III chaeta d3 present. Chaetae associated with trichobotria on abd.II‒III fan-shaped. Chaetotaxy and trichobothrial complex on abd.IV as in Figs 25‒26 View FIGURES 25–27 ; macrochaetae B5, B6, C1, D3, E2, E3, E4, F1, F2 and F3 broader with broad socket, while D2, De3, E1, E4p, E4p2, F3p, Fe4, Fe5, T6 and T7 thinner with smaller socket; macrochaeta F2 above macrochaeta E3; the ratio of distances between macrochaetae C1‒B5 / B5‒B6 1.2‒1.8; accessory chaeta s associated with trichobotrium T2 absent; all chaetae associated with trichobotria on abd.IV (D1, a, m, pe, pi) fan-shaped; three elongate sensillae (S-chaetae) and five slightly ciliated posterior macrochaetae also present. Dorsal chaetotaxy of abd.V as in Fig. 27 View FIGURES 25–27 .
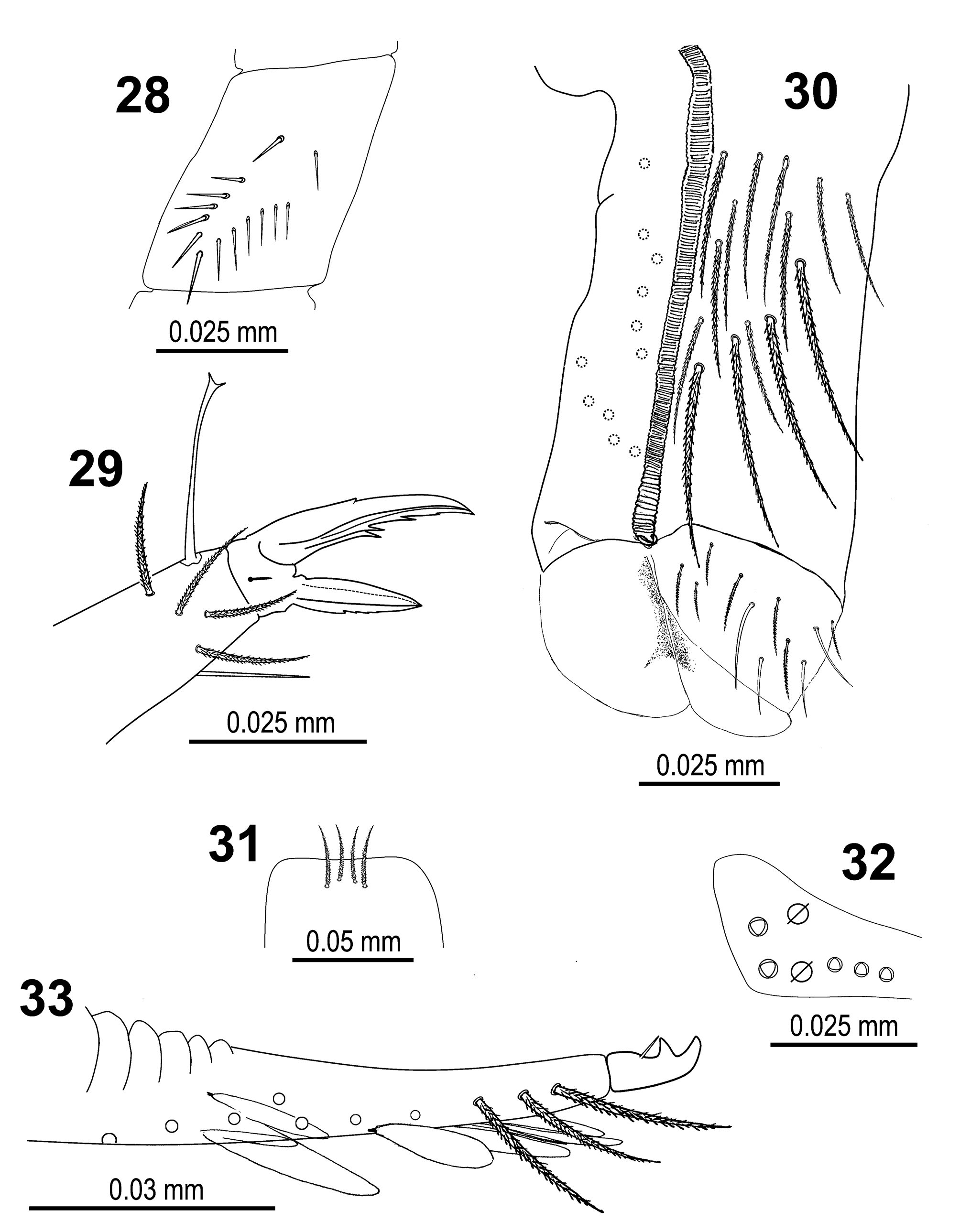
Trochanteral organ with 11‒14 smooth spiny chaetae forming a V shape pattern ( Fig. 28 View FIGURES 28–33 ). Unguis and unguiculus as in Fig. 29 View FIGURES 28–33 ; unguis with paired basal teeth at 52% from the inner edge, one sub-equal median tooth at 70%, and a tiny apical tooth 84% from the inner edge, respectively; a small external tooth also present; unguiculus lanceolate, with 3‒7 denticles along outer edge or with outer edge smooth (in five out of the 19 specimens studied); tibiotarsal tenent hair spatulate, as long as claw; ratio of supraempodial chaeta (smooth chaeta on tibiotarsus III opposite to tenent hair) / unguiculus is around 0.8.
Ventral tube ( Fig. 30 View FIGURES 28–33 ) with a maximum of 14+14 ciliated chaetae on the anterior side and 11+11 weakly ciliated chaetae on the posterior side; lateral flap with a maximum of 10 laterodistal chaetae (7 ciliated and 3 smooth).
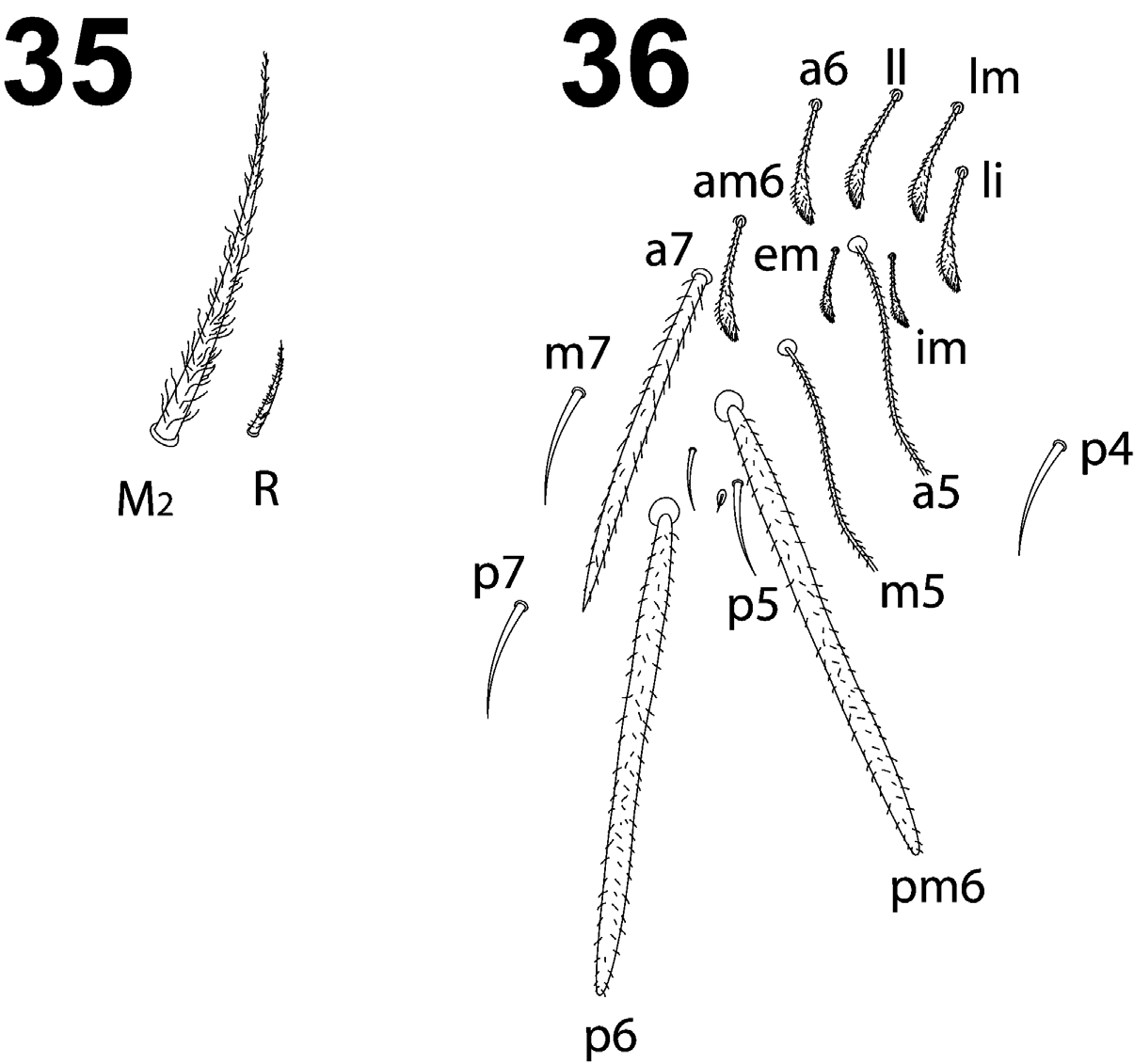
Manubrium with 2+2 ciliated apical chaetae on anterior side ( Fig. 31 View FIGURES 28–33 ); manubrial plate with 2 inner chaetae and 3 chaetae outer to the 2 pseudopores ( Fig. 32 View FIGURES 28–33 ). Dental tubercle absent. Mucro as in Fig. 33 View FIGURES 28–33 . Ratio manubrium/ dens/mucro as 16:18:1.
Variability. On one specimen abd.IV accessory chaeta m doubled (bilateral variability).
Ecology and distribution. The specimens of L. florae sp. nov. were found in moss on alder logs (dead lying trees) and, in low abundance, also in moss on buttressed roots of living alder trees ( Fig. 34 View FIGURE 34 ). As L. florae sp. nov was lacking in the soil samples taken from the type locality, the new species can be considered as a typical bryophilous, hygrophil species.
Discussion. Lepidocyrtus florae sp. nov. shares dorsal trunk macrochaetotaxy 00/0101+2 with seven European species of the genus, namely L. pseudosinelloides Gisin, 1967 (in: Gisin 1967a); L. serbicus Denis, 1933 ; L. tomosvaryi Winkler & Traser, 2012 ; L. isabelleae Winkler, 2017 ; L. arrabonicus Traser, 2000 ; L. weidneri Hüther, 1971 and L. pallidus Reuter, 1890 ( Tab. 1). Also the new species share trunk macrochaetotaxy with the species L. szeptyckii Rusek, 1985 (species with dark violet color, originally described from Siberia and also found in Slovakia, see Kováč et al. 2001), but can be clearly differentiated from it because L. szeptyckii has scales on ant.I‒II, all segments of the three pairs of legs, and posterior face of the manubrium ( Rusek 1985), while L. florae sp. nov. not. Dorsal head chaetotaxy of L. szeptyckii is confusing in the original description (Rusek 1 985, Fig. 2A View FIGURES 1–2 ) and it is not possible to clearly establish which dorsal macrochaetae are present. Rusek only unambiguously described that macrochaetae M2 and S3 were absent. Besides the characters mentioned, L. szeptyckii and L. florae sp. nov. also differentiate by the morphology of apical labral chaetae (divided in L. szeptyckii and pointed in L. florae sp. nov.) and labral papillae (with expansions in L. szeptyckii and rounded in L. florae sp. nov.).
By the absence of dorsal cephalic macrochaetae M2 and presence of macrochaeta S3, the new species shows close affinities with L. pallidus and L. weidneri . By the presence of macrochaeta A3, L. florae sp. nov. clearly differs from both L. pallidus and L. weidneri . Further differences include the different body colour pattern (uniformly dark blue in L. florae sp. nov. and diffuse blue pigment in L. pallidus and L. weidneri ), and the proportionally larger eyes G and H, the absence of abd.II chaeta a2p and the presence of abd.III chaeta d 3 in the new species. From L. weidneri , the new species also differs by the morphology and position of chaeta F2 on abd.IV, which is a smooth mesochaeta in L. weidneri positioned above macrochaeta E2, while in L. florae sp. nov. it is a ciliated macrochaeta below E2. From L. pallidus , the new species can be differentiated also by the ciliated labial chaeta R (in L. pallidus , it is a vestigial, smooth spine-like chaeta).
The new species shows similarities in most features with L. arrabonicus ( Tab. 1), but can be clearly differentiated from it by the presence of dorsal head macrochaeta S3. All the remaining species ( L. pseudosinelloides , L. serbicus , L. tomosvaryi and L. isabelleae ) with dorsal trunk macrochaetae formula 00/ 0101+2 bear dorsal cephalic macrochaeta M2 and consequently are clearly separated from L. florae sp. nov. ( Tab. 1). It should be noted that, in the genus Lepidocyrtus , the only difference in the presence/absence of one dorsal macrochaeta (in the head or in the body) is a robust character for specific differentiation, and no variability in dorsal head macrochetotaxy has been described so far for any Lepidocyrtus species.
Key to the European Lepidocyrtus species without scales on antennae, legs (beyond coxae and femur III) and posterior face of the manubrium, and with dorsal trunk macrochaetotaxy 00/0101+2
The European Lepidocyrtus species without scales on antennae, legs (beyond coxae and femur III) and posterior face of the manubrium, and with dorsal trunk macrochaetotaxy formula 00/0101+2 belong to the pallidus – serbicus group as defined by Winkler & Traser (2012). Subsequently, Winkler (2017) subdivided this group in three, based on dorsal cephalic chaetotaxy. The new data provided by the present work shed light on the actual dorsal cephalic chaetotaxy of the species from pallidus – serbicus group. With this new information, and irrespective of the number of groups, the following species key help to differentiate between all European species described with dorsal trunk macrochaetotaxy as 00/0101+2. In the key the species L. szeptyckii is expressly omitted. The presence of scales on antennae, legs and posterior face of the manubrium clearly separate this species from the species of pallidus – serbicus group. Also we have some preliminar evidence (unpublished data) suggesting that in L. szeptyckii the dorsal macrochaetae of abd.IV correspond to chaetae C1, B4, B5 (as broad ciliated macrochaetae), and B6 (as thin ciliated macrochaeta with reduced socket); so actually the abd.IV dorsal macrochaetae formula of this species would have to be 1+3. In the species of pallidus – serbicus group these chaetae correspond to C1, B5 and B6 (as broad ciliated macrochetae), with B4 as smooth mesochaeta (abd.IV dorsal macrochaetae formula 1+2). The species L. fimicolus is also omitted in the key because it has not yet been detected in Europe.
1 Cephalic macrochaetae S3 and M2 absent ( Fig.1 View FIGURES 1–2 )......... L. arrabonicus Traser, 2000 ( Hungary, Italy, Moldova, Rumania)
- Cephalic macrochaeta S3 present, M2 present ( Fig. 2 View FIGURES 1–2 ) or absent ( Figs 5 View FIGURES 4–5 and 18 View FIGURES 18–19 )..................................... 2
2 Cephalic macrochaeta M2 absent ( Figs 5 View FIGURES 4–5 and 18 View FIGURES 18–19 )............................................................ 3
- Cephalic macrochaetae M2 present ( Fig. 2 View FIGURES 1–2 )................................................................ 5
3 Cephalic macrochaeta A3 present ( Fig.18 View FIGURES 18–19 ); abd.II chaeta a2p absent ( Fig. 23 View FIGURES 23–24 )................ L. florae sp. nov. ( Hungary)
- Cephalic macrochaeta A3 absent ( Fig. 5 View FIGURES 4–5 ); abd.II chaeta a2p present............................................. 4
4 Labial chaeta r vestigial , spine-like microchaeta ( Fig. 4 View FIGURES 4–5 )............................................................................................ L. pallidus Reuter, 1890 ( Belgium, Britain, Finland, Latvia, Norway, Portugal)
- Labial chaeta R short, but ciliated and well developed ( Fig. 35 View FIGURES 35–36 ).................... L. weidneri Hüther, 1971 ( Germany)
5 Abd.II chaeta a2p absent, m3e present (as in Fig. 23 View FIGURES 23–24 )........... L. pseudosinelloides Gisin, 1967 (in: Gisin 1967a) ( Spain)
- Abd.II chaeta a2p present, m3e absent.................................................................... 6
6 Dental tubercle present.......................................... L. tomosvaryi Winkler & Traser, 2012 ( Hungary)
– Dental tubercle absent................................................................................. 7
7 Eyes G and H strongly reduced in size (eventually absent), abd.III a7 a mesochaeta (as in Fig. 24 View FIGURES 23–24 )...................................................... L. serbicus Denis, 1933 ( Greece: Crete, Poland, Hungary, Romania, Serbia, Slovakia)
- Eyes G and H only slightly reduced in size, abd.III a7 a macrochaeta ( Fig. 36 View FIGURES 35–36 )................... L. isabelleae ( Hungary)






| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |