
Cylindera (Eugrapha) contorta (Fleutiaux)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4664.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:A9079148-0462-45E2-8C7F-026CC7B8B196 |
|
DOI |
https://doi.org/10.5281/zenodo.5611519 |
|
persistent identifier |
https://treatment.plazi.org/id/56640051-FFE3-D908-FF3F-FC5C7FA0FC6D |
|
treatment provided by |
Plazi |
|
scientific name |
Cylindera (Eugrapha) contorta |
| status |
|
Cylindera (Eugrapha) contorta View in CoL
Description of third instar larva. Upper part of head brown or dark brown, with weak bronze luster (sometimes greenish hue), lower part of head light brown. Other appendages light brown, except for darkened apical part of mandibles. Maximum width of head at level of stemma I. Setae pale, thin, long and acute apically; some setae near stemmata I-II slightly obtuse. Epicranial suture distinct, but short.
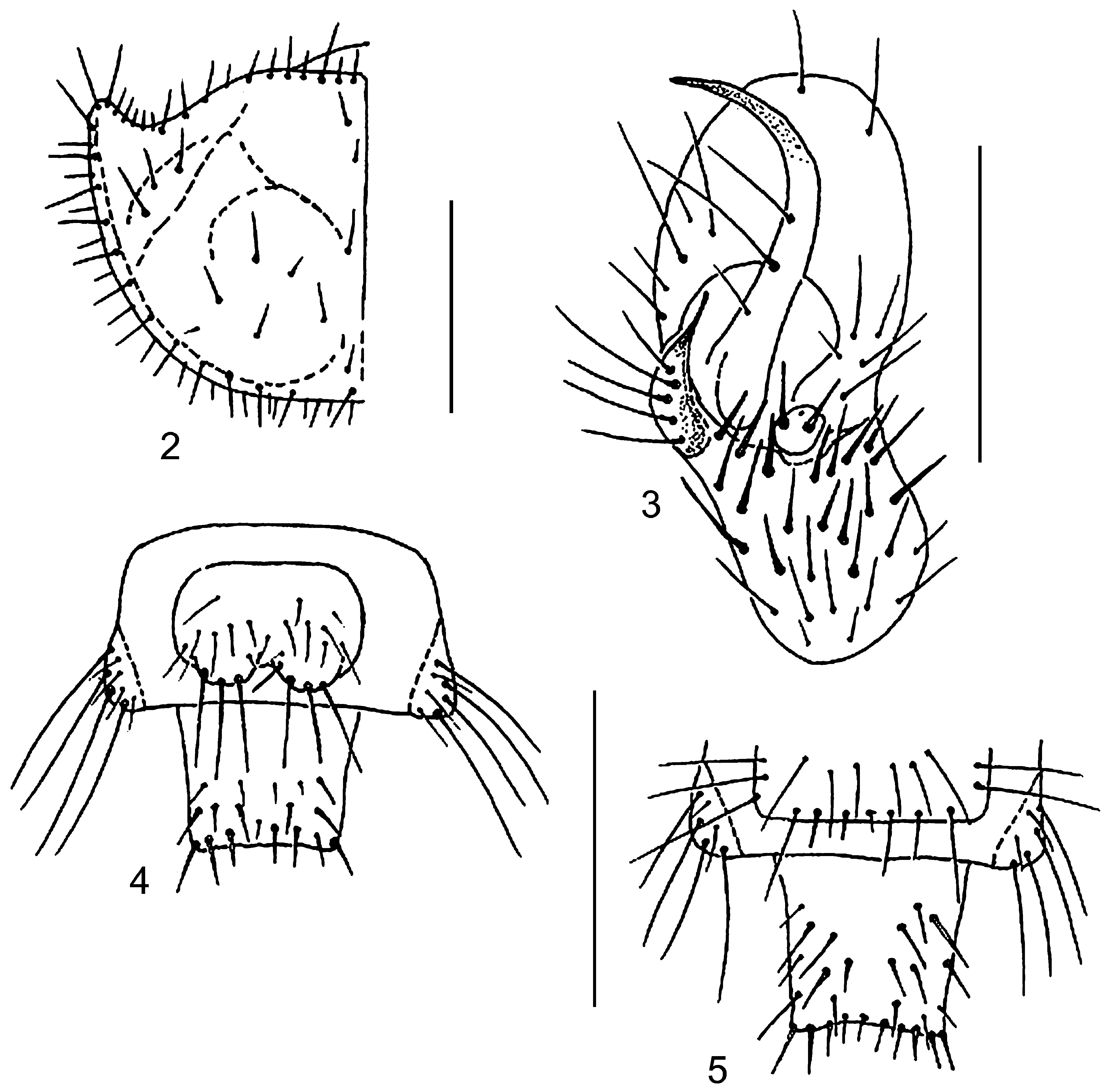
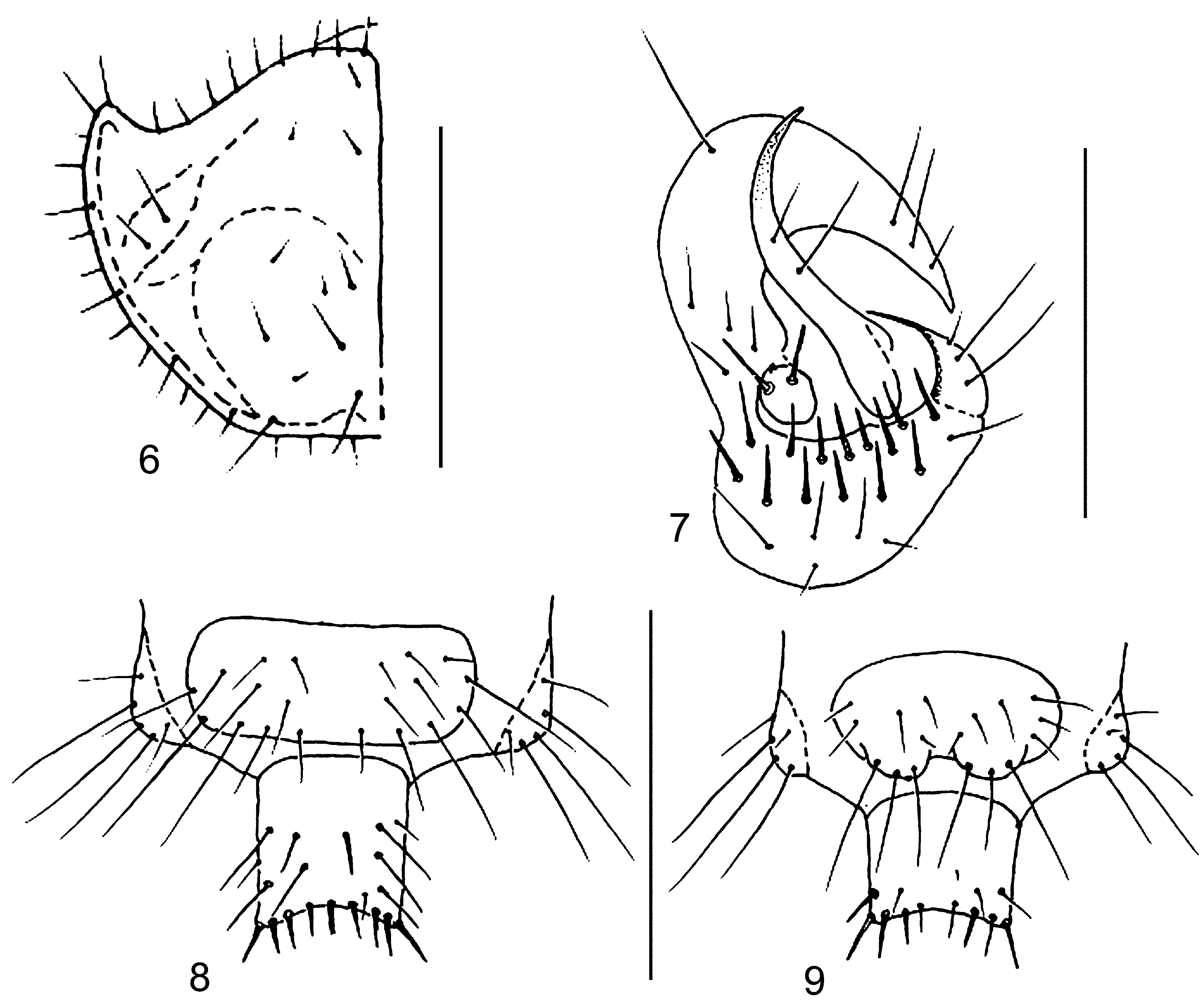
Disk of PN dark brown with weak coppery luster. Apices of PNa and narrow marginal sides of PN lighter. Swellings of PNa and callous elevations on disk moderately distinct ( Fig. 2 View FIGURES 2–5 ). PNa widened with slightly rounded apices directed somewhat outwards and shorter than almost even anterior margin of PN. Setae pale with a reddish hue, most of them thin and long. PN1 with 12–16 setae, and 5–6 setae along PNm, ridge of PNa with 2–3 setae ( Fig. 2 View FIGURES 2–5 ).
Sclerotized areas of abdomen indistinct, gray with reddish hue. T 3 almost square or oval with 6–9 reddish setae. Type II hypopleuron (two large sclerites). CT5 and AT5 light brown, almost semicircular, widely fused on inner margin and partly in contact on external margin ( Fig. 3 View FIGURES 2–5 ). Sclerotization of AT5 covers inner part of IH ( Fig. 3 View FIGURES 2–5 ). LCT5 with 4–5 long setae. CT5 with 20–25 stout setae (similar to those of IH) and some thin small setae on posterior margin. Most setae of AT5 located on its lower (external and inner) part ( Fig. 3 View FIGURES 2–5 ). MH with 3 (rarely 2) setae at the middle. MH slender, very long and strongly curved above the middle. Tips of MH traversing anterior margin of AT5 and distinctly directed outward. MH almost 5–6 times as long as wide at base. IH short, with two long stout setae and directed almost upwards. Central spine of IH absent or point-shaped ( Fig. 3 View FIGURES 2–5 ). IH 5.6–6.0 times shorter than MH. TE9 with 8 setae (4 of them at the middle shortened) ( Fig. 5 View FIGURES 2–5 ). EU9 with 2 groups of 3 long setae each ( Fig 4 View FIGURES 2–5 ). PY (except apex) with 10–17 setae dorsally and 10–12 thinner setae ventrally. Tip of PY with 12–16 bristles, setae on dorsal side shorter.
Description of second instar larva. Upper part of head brown, with weak bronze-copper luster; lower part of head light brown. Setae pale or reddish transparent; most setae thin and acute, some obtuse apically. Epicranial suture distinct, but very short.
Disk of PN and head of same coloration. Apices of PNa widened and directed forwards. Anterior margin of PN greatly elongate, much longer than apices of PNa ( Fig. 6 View FIGURES 6–9 ). Setae of PN1 pale or light brown. Marginal setae reddish, transparent; most of them thin and long, acute, some setae obtuse. PN1 with 10–14 setae, and 4–5 setae along PNm, ridge of PNa with 2–3 setae ( Fig. 6 View FIGURES 6–9 ).
Sclerotized areas of abdomen slight distinct, gray. T 3 almost square, with 5–7 reddish setae. CT5 and AT5 light brown, widely fused on inner margin and divided on external margin. Sclerotization AT5 covers inner part of IH too ( Fig. 7 View FIGURES 6–9 ). LCT5 with 2–3 long setae. CT5 with 12–16 stout setae and some thin small setae on posterior margin. Most setae of AT5 located on lower part ( Fig. 7 View FIGURES 6–9 ). MH with 2 (rarely 3) setae on middle. MH slender, long and distinctly curved above of upper third. Tips of MH reaching (slight traversing) anterior margin of AT5 and directed outward. Length of MH almost in 4.5–5.0 times more width at base. Central spine of IH present but very small ( Fig. 7 View FIGURES 6–9 ). IH length in 5 times less those MH. TE9 with 8 setae (4 of them on middle shortened) ( Fig. 8 View FIGURES 6–9 ). EU9 with 2 groups of 3 long setae each ( Fig. 9 View FIGURES 6–9 ). PY (except apex) with 10–12 setae dorsally and 5–7 ventrally. Tip of PY with 12–14 setae.
Description of first instar larva. Coloration of head and pronotum the same as in second and third instar larvae, but without luster. Setae pale; most setae thin and acute, some obtuse apically. Epicranial suture present, but indistinct.
PNa more widened and directed outwards; its apices rounded. Anterior margin of PN much longer than apices of PNa and middle region very distinctly concave ( Fig. 10 View FIGURES 10–13 ). All setae of PN transparent pale. Most of them thin and long, acute; some marginal setae obtuse. PN1 with 5–6 setae, 3–4 setae located along PNm. PNa without setae ( Fig. 10 View FIGURES 10–13 ).
Sclerotized areas of abdomen indistinct, grayish. T 3 almost square with 3 reddish setae. CT5 and AT5 light brown, indistinctly contacting at inner margin. Lower parts AT5 and CTL5 more sclerotized; AT5 with 2 setae ( Fig. 11 View FIGURES 10–13 ). CT5 without setae. LCT5 with one long seta. MH with one long seta above the middle. MH distinctly narrowed and curved above the middle. Tips of MH reaching anterior margin of AT5. MH almost 4.0 times as long as wide at base. Central spine of IH present, but very small ( Fig. 11 View FIGURES 10–13 ). IH no more than 3.5 times as long as MH. TE9 with 6 setae ( Fig. 12 View FIGURES 10–13 ). EU9 with 2 groups of 3 long setae each ( Fig. 13 View FIGURES 10–13 ). PY (except apex) with 6 setae dorsally and bare ventrally. Tip of PY with 10 setae.
Distribution and ecological peculiarities. In Southern Ukraine the tiger beetle Cylindera contorta is found sporadically at open areas of the sea coastline: at the outer coasts of the Kinburn, Tendra and Dzharylgach sand spits of the Kherson Region, whose total coastal line is 142 km long ( Fig. 1 View FIGURE 1 ).
Adults and larvae of the species are moderately halophilous, thus concentrated at small narrow saline lagoons, 3–5 m wide and 5–30 m long, less at the zone of sea surf ( Figs. 14–17 View FIGURES 14–17 ).
The life cycle of C. contorta lasts from one to two years, but a two years cycle is more common ( Putchkov & Nitochko, 2016). Oviposition is observed from late spring (May) to mid-summer (July). The overwintering phase is usually third instar larvae, rarely the adults hibernate. Pupation takes place from end of May to June. Adults live from 3 months to 1–2 years, while larvae live up to 2 years. Copulation ( Fig. 19 View FIGURES 19–20 ) is usually observed from first days of June to middle of July. Adults are found at open sandy sea shore areas, occasionally adults are recorded at sandy-shellfish beaches, salt lakes and estuaries ( Fig. 14–15 View FIGURES 14–17 ). Adults are registered from early May to late August, in highest quantities from late May to mid-July. The species is locally distributed, though occasionally abundant. The larvae of first and second instars are recorded at first and second decades of July, and third instar larvae are found from the end of July to middle of August. Larvae prefer only the sandy areas at the littoral zone of quiet bays, estuaries and salt lakes ( Fig. 16–17 View FIGURES 14–17 ), as well as at the depressions with sufficient soil moisture; and larvae discard the sandy areas at zone of sea surf ( Fig. 20 View FIGURES 19–20 ).
Larvae usually aggregated at certain elements of mesorelief ( Fig. 16–17 View FIGURES 14–17 ): small lagoons 3–5 m wide and of various lengths (usually 5-30 m long). Moreover, the burrows made by larvae are found at very limited section of lagoons, along their outer contour, but not in the very moist central part ( Fig. 18 View FIGURES 18 , 20 View FIGURES 19–20 ). Larvae cover the upper part of burrows with soil (sometimes to one third of the total length) when the cold weather, heavy rain or very hot (dry) periods come and remain inactive until conditions improve. Under excessive moisture or habitat destruction caused by storms, the larvae leave the burrows and migrate (usually at night) to more favorable areas, sometimes up to several tens of meters away. Usually larvae can be found only in limited periods of the year: from the end of July to August.
Adults and larvae of Cylindera contorta are often found simultaneously with the Eurasian species Calomera littoralis (Fabricius) . However, these species differ in bionomy and habitat preferences.
Adults and larvae of C. littoralis are numerous throughout the littoral zone (beetles are often recorded far from water): at areas with dense clay soils to sands with varying degree of moisture. The species is common during all warm season, from April-May to September. They usually overwinter as both, adults and third instar larvae. The oviposition is usually prolonged and lasts from the beginning of May to the end of August. Thus, specimens of C. littoralis are developing asynchronously. The depth of larval burrows vary from 15 cm (first and second instar larvae) to 25 cm (third instar larvae).
Adults and larvae of C. contorta occur mostly at sandy coastal areas. Beetles are found from the beginning of May to the end of second decade of August, with maximal abundance from the middle of July to mid-August. They usually overwinter as third instar larvae. Most of first and second instar larvae occur at July, but third instar larvae are recorded since first half of August. Thus, specimens in studied populations of C. contorta are developing synchronously. The depth of larval burrows varies from 8–15 cm (first and second instar larvae) to no more than 20 cm (third instar larvae).
Protection and limiting factors. In Ukraine, over 90% of habitats of C. contorta is covered by the Ramsar sites. These habitats are partially protected at the Black Sea Biosphere Reserve, “Dzharylgachsky” National Nature Park and “Beloberezhie of Svyatoslav” National Nature Park (all in Kherson Region).
A number of factors limit the abundance of C. contorta in Ukraine: small habitat areas, storms that destroy the main biotopes (lagoons and sea spits), flooding of areas with larval burrows. However, the anthropogenic pressure is especially dangerous, namely the urbanization of sea coasts: development of tourist infrastructure with recreation centers and resorts (especially in Dzharylgach island).
Comparative notes. In Cicindelidae the larvae of different instars (including Eugrapha ) are well distinguishable by some qualitative and quantitative characteristics. The most important are the proportions of pronotum, especially its length/width ratio that varies from 1.41–1.53 ( R 1, R 2) to 2.15–2.40 ( R 3) in different larvae of the subgenus (table 2 and abbreviations of the table). Moreover, the larval instars are well identified by the chaetotaxy of different segments. The most important and constant feature of chaetotaxy is the number of setae on the second segment of the galea: one seat for first instar larvae; two setae for second instar larvae; three or four setae for third instar larvae.
In addition, other peculiarities can be used to identify different larval instars of Eugrapha and other Cicindelina:
– no setae on AN1, PNa, and CT 5 in first instar larvae; 4–6, 1–3, and 13–25 setae on AN1, PNa, and CT 5 in second and third instar larvae respectively;
– only 3–4 setae on T 3 in first instar larvae; 5–9 setae on T 3 in second instar larvae; 6–15 setae on T 3 in second and third instar larvae;
– LCT5 with one bristle in first instar larvae; LCT5 usually with 3–5 bristles in second instar larvae; LCT5 with 5–10 bristles in third instar larvae;
– LP1 with only 3 setae in first instar; LP1 usually with 5 setae in second instar; LP1 with 7 setae in third instar larvae.
Our previous research revealed, that all Eugrapha larvae are characterized by some specific features and they are well distinguished from other larvae the genus Cylindera and other genera of the subtribe Cicindelina ( Putchkov, 2001, 2013).
Thus, second and third instar larvae of the genus Eugrapha exhibit following combination of characters:
– central spine of IH absent or very small (short);
– IH very short, usually 4–5 times shorter than MH;
– MH slender and usually strongly curved in upper third, their tips usually reaching anterior margin of AT5 or longer;
– tergites of fifth abdominal segment always fused on inner margin;
– sclerotized areas of abdomen slightly distinct;
– EU9 always with 2 groups with 3 setae;
– anterior margin of PN longer than apices of PNa;
– head and PN no more than 1.8 mm wide in second instar larvae and no more than 2.8 mm wide in third instar larvae.
The first instar larvae of the genus Eugrapha differ in having following combination of characters (in contrast to larvae of other Cicindela s.l.):
– AT5 of the fifth abdominal segment is more sclerotized along posterior inner margin only (as separate areas); – MH long, slightly curved, but distinctly narrowed in the upper third;
– central spine of IH distinct, but short;
– anterior margin of PN strongly convex, distinctly longer than cephalolateral angles;
However, many larval characters of Eugrapha , especially features of chaetotaxy, taken separately appear to be relatively similar or overlapping within the subgenus (table 3). Thus, determination of the species becomes reliable only based on a complex of features: these are given in an updated key for larvae of the subgenus Eugrapha (see below).





| T |
Tavera, Department of Geology and Geophysics |
| MH |
Naturhistorisches Museum, Basel |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |