
Miconia galdamesiae Kriebel & Almeda, 2013
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.134.1.2 |
|
persistent identifier |
https://treatment.plazi.org/id/57230B55-4963-D74E-FF01-76D4FE64F868 |
|
treatment provided by |
Felipe |
|
scientific name |
Miconia galdamesiae Kriebel & Almeda |
| status |
sp. nov. |
Miconia galdamesiae Kriebel & Almeda View in CoL sp. nov.
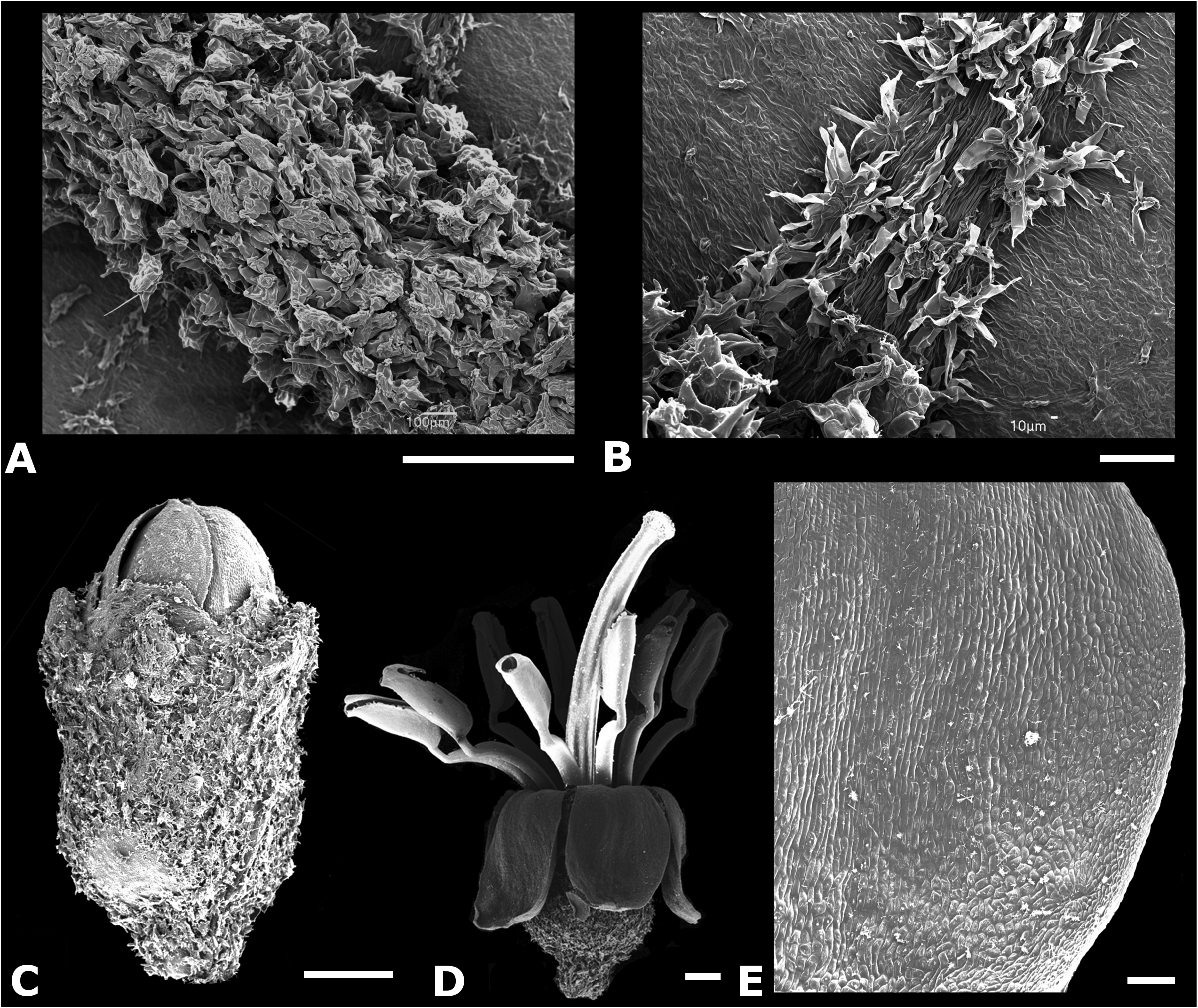
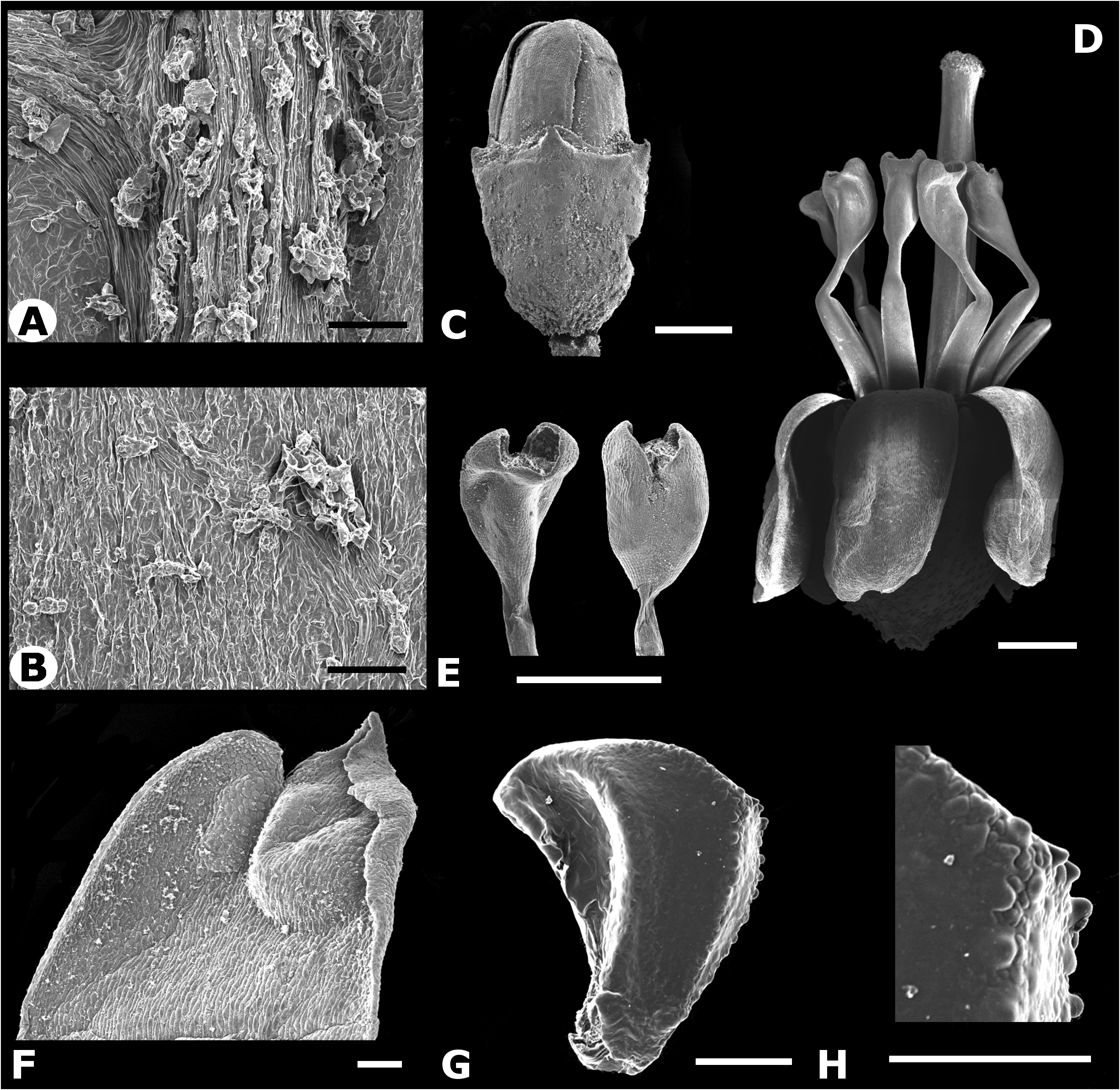
TYPE:— PANAMA. Chiriquí: Reserva Forestal de Fortuna. Sendero atrás de la estación del Smithsonian ( STRI) , 1162 m, 8.734583 N, - 82.240083 W, 19 September 2011, R. Kriebel 5736 & J. Burke (holotype NY!, isotypes INB!, PMA!) . Figs. 1–2 View FIGURE 1 View FIGURE 2 .
Small trees 2.5–7 m tall with young stems orange–brown from the copious indument of asperous–headed hairs, nodal line barely evident. Petioles 0.8–4 cm. Leaf blades 4.5–20 × 2–10.5 cm, 3–5–plinerved, diverging from the midvein 0.5–6 cm above the base usually asymmetrically, elliptic–ovate to ovate, base obtuse to acute and usually oblique, apex acuminate, the margin denticulate, adaxially glabrous except for asperous–headed hairs on the main veins towards the base, somewhat thin and dark green when alive, abaxially densely pubescent on tertiary and higher order veins with asperous–headed orange–brown hairs and glabrous to glabrescent on the actual surface. Inflorescences terminal, lax dichasia branched at or near the base of the inflorescence, 3.7–7 cm long, copiously covered with orange–brown asperous–headed hairs, bracts to 8 mm long, linear oblong, bracteoles 0.5–1 mm long, linear, less pubescent than rest of inflorescence rachis, drying pinkish, flowers clustered at the end of the inflorescence branches. Pedicels essentially absent. Hypanthia campanulate 1.25–1.75 × 1–1.5 mm, densely covered with asperous–headed hairs. Flowers 5– merous. Calyx fused in bud, shortly apiculate and less pubescent than the hypanthium, rupturing at anthesis into irregular, broadly rounded hyaline lobes 0.25–0.75 mm long and 0.5–0.75 mm wide at the base, the exterior calyx teeth 0.25–0.5 mm long, linear oblong, the calyx tube 0.25–0.5 mm long. Petals 1.5–2 x 1–1.5 mm, ovate, white, smooth, reflexed at anthesis, emarginate. Stamens 10, 3– 3.5 mm long, radially arranged around the style; filaments 1.5–2 mm long, geniculate near the apex, translucent white; anthers 1–1.5 × 0.35– 0.65 mm, linear–oblong, somewhat laterally compressed, cream yellow, pores 0.1–0.15 mm wide, truncate to somewhat ventrally inclined. Ovaries 5–locular, half inferior, apex elevated into a low papillose collar. Styles 4.5–4.75 mm long, straight to very slightly curved, distance between the anther apex and the stigma ca. 1 mm; stigmas truncate to capitellate, ca. 0.5 mm wide. Berries described as green–red on one label (McPherson 8410, CAS) but drying purple, 1.7–1.9 × 2.0– 2.2 mm when dry; seeds ovoid and angled, 0.4–0.5 × 0.3–0.4 mm, orange–brown, lateral symmetrical plane ovate to triangular, the highest point toward the chalazal side, antiraphal symmetrical plane ovate–triangular and inconspicuously verruculose on the angles, raphal zone narrowly triangular and extending the length of the seed, expanded into an appendage that covers about 30% of the seed length. Chromosome number: n = 17 (reported as M. aff. friedmaniorum in Almeda 2013).
Distribution— Miconia galdamesiae is known from two cloud forest localities in Panama, one in Chiriquí / Bocas del Toro Provinces and one in Veraguas Province in an elevation range from 860–1350 m.
Phenology— Specimens of Miconia galdamesiae have been collected in flower in January through March, August and September, and with fruit in February and September.
Etymology— It is with great pleasure that we name this species for our colleague, Carmen Galdames, botanist at the Smithsonian Tropical Research Institute who has made many important collections of Melastomataceae and the Panamanian flora in general.
Discussion— Miconia galdamesiae belongs to a group of small tree species that appear to be restricted to the cloud forests of Costa Rica and Panama. Based on molecular phylogenetic data (Kriebel & Michelangeli in prep.) M. galdamesiae groups in a well supported clade that includes M. friedmaniorum , M. papillopetala , and M. pendula Umaña & Almeda (1993: 5) . These species share a similar indument of roughened trichomes, leaves that are typically asymmetrically plinerved with the main veins diverging from the mid vein in alternate fashion, small flowers with a fused calyx that ruptures prior to anthesis, filaments that are geniculate just below the anther thecae, five-locular ovaries, very exserted styles and seeds that are more or less ovoidtriangular in outline and angulate on the antiraphal surface with or without verruculose ornamentation that is restricted to the angle ridges. Miconia galdamesiae can be distinguished from both Miconia pendula and M. friedmaniorum by its ovate petals (vs. linear-oblong petals). Also, M. galdamesiae has sessile flowers in glomerules and exhibits a ‘big bang’ flowering strategy ( Fig. 1 View FIGURE 1 ), whereas the other two species have inflorescences in which few flowers appear to open at a time, and the flowers are pedicellate and not arranged in glomerules. The stamens of M. galdamesiae are declined away from the style at anthesis unlike M. pendula (see Almeda & Umaña 1993, for a drawing of M. pendula ) and M. friedmaniorum ( Figs. 5–6 View FIGURE 5 View FIGURE 6 ) where the stamens are held tightly around the style at anthesis ( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). Miconia pendula also differs in its lanate indument and both M. pendula and M. friedmaniorum differ from M. galdamesiae in their deflexed inflorescences. Continuing botanical exploration in southern Central America suggests that M. friedmaniorum and M. pendula are restricted to Costa Rica as suspected when they were described whereas M. galdamesiae and M. papillopetala are endemic to Panama. Miconia galdamesiae is also similar to and grows sympatrically with M. papillopetala on Cerro Tute (= Cerro Mariposa) in Veraguas Province. The two Panamanian species can be distinguished by petal color and indument details on upper cauline internodes, the inflorescence axes and the leaves (see Table 1 and Figs. 3 View FIGURE 3 & 4 View FIGURE 4 ). See discussion under M. papillopetala below for further comments.
Another close relative of the above discussed taxa is Miconia brenesii which has a broader distribution that may ultimately extend to Panama. This species can be distinguished from M. galdamesiae by its young branches that appear glabrous or with very short roughened hairs, pedicellate, unclustered flowers, a calyx that is not fused prior to anthesis and very broad anther pores that are typically bent backwards toward the dorsal side of the anther ( Figs. 7 View FIGURE 7 & 8 View FIGURE 8 ).
Representative Specimens Examined— PANAMÁ. Bocas del Toro: Fortuna Dam Area , along continental divide trail bordering Chiriquí Province, 1200–1300 m, 08º45’04”N, 82º15’04”W, 10 March 1988, F GoogleMaps . Almeda et al. 6059 ( BM, BR, CAS, GH, MEXU, MO, NY, PMA, P, US); Fortuna Dam region , near trail along continental divide, 11 February 1986, 1250– 1300 m, ca. 8º45’N, 82º15’W, G GoogleMaps . McPherson 8410 ( CAS, MO, PMA); Vicinity of Fortuna Dam, along continental divide trail west of highway, 1250 m, 08°45’N, 82°15’W, 5 September 1987, McPherson 11636 ( CAS, MO, PMA). Coclé: Cerro Tigrero , 1000– 1350 m, 8°37’33”N, 80°41’18”W, 26–28 September 2001, J GoogleMaps . Mendieta 17-466 ( CAS, PMA). Veraguas: Trail to Reserva Biológica Serrania de Tute and the summit of Cerro Tute about 0.7 km beyond the Escuela Agricola Rio Piedra just outside Santa Fe , 18 February 1996, 860– 1300 m, F . Almeda et al. 7618 ( CAS, INB, MO, NY, PMA, US); Vicinity of Escuela Agricola, Alto de Piedra near Santa Fe. 3 mi beyond the fork in the road near the school, toward Atlantic slope along trail to top of Cerro Tute , 26 January 1980, 3400–3800 ft., T . Antonio 3501 ( CAS, MO, PMA); puente sobre río Los Valles, cerca de la hidrolectrica Estrella Los Valles , 28 August 1982, M . D. Correa et al. 4341 ( CAS, PMA); Cerro Tute ridge up from former Escuela Agricola, Santa Fe , 1000–1300 m, 8°05’W, 8°35’N, 20 February 1983, C GoogleMaps . Hamilton & R . Dressler 3073 ( CAS, EAP, INB, MEXU, MO, PMA) .






| STRI |
Smithsonian Tropical Research Institute |
| R |
Departamento de Geologia, Universidad de Chile |
| J |
University of the Witwatersrand |
| NY |
William and Lynda Steere Herbarium of the New York Botanical Garden |
| INB |
Instituto Nacional de Biodiversidad |
| PMA |
Provincial Museum of Alberta |
| CAS |
California Academy of Sciences |
| F |
Field Museum of Natural History, Botany Department |
| BM |
Bristol Museum |
| BR |
Embrapa Agrobiology Diazothrophic Microbial Culture Collection |
| GH |
Harvard University - Gray Herbarium |
| MEXU |
Universidad Nacional Autónoma de México |
| MO |
Missouri Botanical Garden |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| T |
Tavera, Department of Geology and Geophysics |
| M |
Botanische Staatssammlung München |
| C |
University of Copenhagen |
| EAP |
Escuela Agrícola Panamericana |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |