
Protoselene ashtoni, Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee, 2014
|
publication ID |
https://doi.org/10.1206/3797.1 |
|
persistent identifier |
https://treatment.plazi.org/id/587387C9-142F-5170-FE57-1B72FB52FB59 |
|
treatment provided by |
Carolina |
|
scientific name |
Protoselene ashtoni |
| status |
sp. nov. |
Protoselene ashtoni , new species
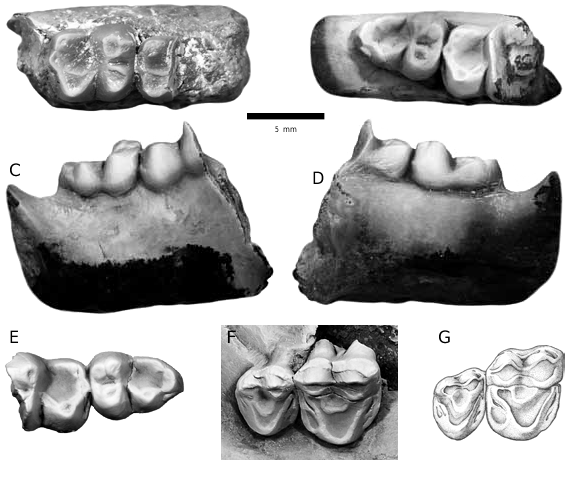
HOLOTYPE: RAM 9047 , left dentary fragment with a partially erupted p4 and m1 from RAM locality V200702, member 4a, Goler Formation , California (fig. 7A–C).
ETYMOLOGY: Named in honor of R. Larry Ashton, Jr., for his support of the research programs of the Raymond M. Alf Museum of Paleontology.
DIAGNOSIS: Protoconid of p4 highly inflated, significantly more than other known p4s of Protoselene , p4 lacking metaconid and anterior basal cusp in contrast to P. opisthacus , paraconid of m1 more lingual than P. opisthacus or P. novissimus , relative length and width of m1 talonid shorter and wider than P. novissimus , and m1 larger than P. griphus .
DESCRIPTION: The distinctive features of the p4 of RAM 9047 are its greatly swollen protoconid and the lack of both an anterior basal cusp and metaconid (fig. 7A–C). On the anterior face of the protoconid is a small, but distinct ridge, that defects posterolabially as it traces the midline of the crown and extends to the apex. A well-developed cristid obliqua extends to the posterior edge of the p4, where it defects lingually and lessens in height. Thus, a small talonid basin is present that opens lingually. Talonid enamel is rugose, in contrast to the faintly crenulated protoconid enamel.
The m1 of RAM 9047 has a low, lingually positioned paraconid connected by a welldeveloped paracristid with a large protoconid. The metaconid and protoconid are subequal in size and height. The talonid basin of m1 opens lingually and is deep, and broad. The cristid obliqua extends posteriorly to the hypoconid, which is larger than the entoconid and the slightly smaller hypoconulid, which are closely appressed. A well-developed precingulid is present anterior to the trigonid, as is a small postcingulid posterior to the hypoconid. The p4 of RAM 9047 is 5.99 mm in length and 4.08 mm in width, and the m1, 5.63 mm in length, with trigonid and talonid widths of 4.13 mm and 4.33 mm respectively.
DISCUSSION AND COMPARISIONS: Protoselene is a relatively rare component in Torrejonian and Tiffanian mammalian faunas from western North America. Found at many Torrejonian localities in the San Juan Basin of New Mexico ( Matthew, 1937; Williamson, 1996), P. opisthacus is the best-known species. Another Torrejonian species is Protoselene (“ Dracoclaenus ”) griphus known from a few specimens collected at the early Torrejonian Dragon Canyon site in central Utah ( Gazin 1939, 1941; Robison, 1986) and a single P4 from the lowest Torrejonian part of Kimbeto Wash in the San Juan Basin ( Williamson, 1996). The third species is P. novissimus from early Tiffanian strata in the Bison Basin of southwest Wyoming ( Gazin, 1956). A few other specimens of Protoselene have been identified from Tiffanian strata, including fragmentary teeth of P. cf. P. opisthacus from the Torrejonian–Tiffanian overlap zone and early Tiffanian of the Hanna Formation of southern Wyoming ( Higgins, 2003), and a dentary with m1–3 of P. opisthacus from Ray’s Bonebed (middle Tiffanian) from the Big Bend area of Texas ( Schiebout, 1974).
The massive and swollen p4 protoconid of P. ashtoni is its most distinctive feature and separates it from P. opisthacus whose protoconid is only slightly inflated. Also, the p4 of P. opisthacus usually has an anterior basal cusp and a small metaconid, features absent in RAM 9047. The p4 is not known for either P. griphus or P. novissimus , so comparisons of these species to P. ashtoni are limited to the m1. The only known m1 of P. novissimus (USNM 21023) is an isolated tooth ( Gazin, 1956) with a relatively longer and narrower talonid than the m1 of P. ashtoni . Also, the m1 paraconid of P. ashtoni is positioned more lingually than in either P. novissimus or P. opisthacus . The only known m1 of P. griphus (USNM 15773) has a lingually positioned paraconid like the m1 of RAM 9047, but the m1 of P. ashtoni is larger than the m1 of P. griphus as the length and width of USNM 15773 are 5.3 mm and 4.0 mm, compared to 5.63 mm and 4.33 mm for RAM 9047. Also, a slight accessory cusp is present on the anterior dentary fragment with m2 and talonid of
FIGURE 7. Dentition of the holotype of Protoselene ashtoni , m1 ( fig. 8A View FIGURE 8 ), and RAM 6926, right maxnew species, RAM 9047, left dentary fragment with m1 and illa fragment with M2–3 (referral ques-
partially erupted p4 from RAM locality V200702; A. occlusal view; B. lingual view; C. labial view. tionable) ( fig. 8F–G View FIGURE 8 ) from RAM locality V200001, member 4a.
ETYMOLOGY: Named in honor of the late Steven Walsh, whose enthusiastic leadership of Goler Formation screenwashing efforts significantly increased the sample of Tiffanian vertebrates from southern California.
DIAGNOSIS: Significantly larger than all other described species of Promioclaenus .
DESCRIPTION: Lower premolars of Promioclaenus walshi are unknown. The m1 of the holotype ( RAM 9098 ) is broken, with only the labial half present, which preserves part of the talonid basin and a large hypoconid and protoconid. A complete but worn m1 talonid is present in RAM 6724 ( fig. 8A View FIGURE 8 ) and the hypoconid is large and the talonid basin is well developed and opens lingually through a deep talonid notch. A distinct entoconid and hypoconulid are present, with the hypoconulid slightly smaller than the entoconid and placed medially, while the entoconid is positioned on the posterolingual tooth margin. The hypoconulid is positioned closer to the entoconid than the hypoconid and a small postcingulid is developed between the hypoconid and hypoconulid .
RAM 6724 has a slightly worn, but well preserved, quadrate shaped m2 ( fig. 8A View FIGURE 8 ). The trigonid is anteroposteriorly short because the large metaconid and small paraconid are closely appressed and the paraconid is slightly shifted labially. The paracristid is faintly visible while the protocristid is distinct. The talonid of m2 is very similar to m1, with a large hypoconid and smaller hypoconulid and entoconid, with the entoconid the larger of the two. The partial m2s of the holotype ( fig. 8B–E View FIGURE 8 ) have the same features as the complete m2 of RAM 6724 .
Left and right m3s of the holotype are identical in size and morphology ( fig. 8B–E View FIGURE 8 ). The protoconid is the largest trigonid cusp and the paraconid and metaconid are subequal in size, with a slight labial shift of the paraconid. The paracristid is more distinct in m3 than in m2. The m3 talonid has a large hypoconid, broad hypoconulid and a smaller ridgelike entoconid positioned directly adjacent to the hypoconulid, separated by a slight constriction in the talonid wall. The talonid basin is narrow, but deep, and opens lingually through a small talonid notch.
A maxilla fragment with right M2–3 (RAM 6926) was found in a concretion at the same site as RAM 6724 and may represent the upper dentition of Promioclaenus walshi . The maxilla fragment is crushed making it difficult to photograph M2–M 3 in occlusal view ( fig. 8F View FIGURE 8 ), so a drawing is provided ( fig. 8G View FIGURE 8 ). The M2 is quadrate in outline and has moderate to heavy wear on all major cusps. The protocone is the largest cusp and the paracone is slightly larger than the metacone. A centracrista wear facet connects the metacone and paracone, as do preparacrista and postprotocrista wear facets connect the protocone with the paraconule and metaconule. The precingulum and postcingulum are both present, and a well-developed hypocone is positioned on the postcingulum. Before wear, it appears that the apex of the hypocone was positioned directly posterior to the apex of the protocone or slightly labial to it. The lingual cingulum is not developed across the lingual face of the protocone and the labial cingulum is developed only adjacent to the ectoflexus and paracone. A minute parastyle and weak ectoflexus are present, but the metastyle is absent.
The M3 of RAM 6926 has a rectangular shape and is significantly smaller than M2. The protocone is the largest cusp, with the metacone and paracone subequal in size. The metacone of M3 is shifted lingually in comparison to its position in M2. The paraconule and metaconule are also relatively smaller in M3 compared to M2. A distinct hypocone is not evident on the postcingulum of M3.
DISCUSSION AND COMPARISIONS: The lower dentition of Promioclaenus walshi is essentially a larger version of the morphology seen in P. lemuroides from the Nacimiento Formation of
A B
New Mexico described by Matthew (1897, 1937). Compared to the holotype of P. lemuroides , lower molars of P. walshi are consistently larger ( table 9). In addition to the holotype, 12 dentary and three maxillary fragments of P. lemuroides at the AMNH, were also measured to document the range in dimensions of premolars and molars of P. lemuroides . These results also indicate that P. walshi is significantly larger than P. lemuroides , particularly in lower molar width ( table 9). Promioclaenus walshi is also larger than P. pipiringosi , a poorly known species described by Gazin (1956) from early to middle Tiffanian strata in the Bison Basin of Wyoming. The holotype of P. pipiringosi (USNM 20571) is a partial dentary whose molars are similar in morphology and size to those of P. lemuroides ( table 9). However, Gazin (1956) also referred a dentary fragment with p2–4 (USNM 21021) from the type locality to P. pipiringosi . If this referral is correct, P. pipiringosi has smaller and less inflated premolars than P. lemuroides ( table 9). The lower premolars of P.walshi are unknown, so no comparison can be made with P. pipiringosi or P. lemuroides . Promioclaenus acolytus is the only other species of Promioclaenus . The size of its lower dentition, based on the large sample from Swain Quarry ( Rigby, 1980: table 46), is much smaller than P. walshi ( table 9).
A second species of Promioclaenus , that is larger than P. pipiringosi , may occur in the Bison Basin. A fragmentary dentary with the posterior portion of m2 and part of the trigonid and roots of m3 (USNM 21020) was noted as possibly representing Litaletes or another species of Promioclaenus ( Gazin, 1956) . The talonid of the m 2 in USMN 21020 has an entoconid, hypoconulid, and hypoconid that have about the same relative size and positioning as those cusps in Promioclaenus . The size of the m 2 in USNM 21020 (based on a photograph with scale) shows a length and width of approximately 4.75 mm and 4.00 mm, respectively, 0.3–0.4 mm larger than P. pipiringosi and 0.4–0.5 mm smaller than P. walshi ( table 9). Gazin (1956) surmised that if USMN 21020 represented Promioclaenus , it was too large to be P. pipiringosi . USNM 21020 is too fragmentary for detailed comparison to P. walshi , but its smaller size suggests it is not the Goler Formation species.
A partial maxilla with M2–3 (RAM 6926) from RAM locality V200001, the same site that yielded a partial dentary of P. walshi (RAM 6724) could represent the upper dentition of P. walshi . RAM 6926 was compared to the M2–3 of P. lemuroides and similarities include the relative size and position of the protocone, metacone, paracone, and conules, and cingulum development. However, there are significant differences, as RAM 6926 has a more quadrate shaped M2 (like the M1 of P. lemuroides ), a relatively larger M2 hypocone, an M3 closer in size to M2 compared to the significantly smaller M3 relative to M 2 in P. lemuroides , and an anteroposteriorly broad, rectangular M3, compared to the narrow M 3 in P. lemuroides . Thus, M2–M3 of P. lemuroides and RAM 6926 have the same cusp construction and arrangement, but proportional differences in certain cusps and tooth shape cast doubt on whether RAM 6926 represents P. walshi , although these differences might represent characters that distinguish P. walshi from other species of Promioclaenus .
A maxilla with heavily worn M2–3 (USNM 21022) from the Bison Basin that was tentatively referred to P. pipiringosi ( Gazin, 1956) has some bearing on this issue. The M2 of USNM 21022 is complete and has a distinct rectangular shape, measuring about 3.2 mm in length and 5.0 mm in width. Although heavily worn, the wear facets indicate that the paracone of USNM 21022 was slightly larger than the metacone and a distinct hypocone was present on the posterior cingulum. Thus, two discernible features of the M2 of USNM 21022 resemble features present in the M2 of RAM 6926, but the shape of the M2 of USNM 21022 is rectangular like P. lemuroides , unlike the quadrate-shaped M2 of RAM 6926. Also, the M3 of USNM 21022 is nearly as large as the M2, more so than in either P. lemuroides or P. walshi . It is uncertain whether USNM 21022 even represents P. pipiringosi , but if it does, then upper molar morphology of Promioclaenus would exhibit significant variation. The two molars preserved in RAM 6926 are about the correct size to represent the upper dentition of P.walshi . But based on differences in upper molar morphology evident in RAM 6926, P. lemuroides , and USNM 21022 ( P. pipiringosi ?), referral of RAM 6926 to P. walshi is tentative.
Tooth P.walshi P. pipiringosi P. lemuroides P. lemuroides P. acolytus Site Holotype
L — 2.7 3.65–3.70 3.90 —
P2
W — 1.6 1.90–1.97 2.09 —
L — 3.3 4.50–4.79 4.55 3.60–3.90
P3
W — 2.1 2.52–2.84 2.83 2.10–2.40
L — 3.8 4.55–4.85 4.62–4.72 3.45–4.35
P4
W — 2.8 3.16–3.47 3.36–3.45 2.55–3.15
L 5.10 4.1 4.07–4.80 4.19–4.36 3.15–4.05
M 1 WTri — — 3.23–4.22 3.67–3.87 2.40–3.45 WTal 4.05 3.4 3.35–4.01 3.77–3.78 2.55–3.45
L 4.96*–5.50 4.4 4.23–4.73 4.26–4.38 3.45–4.50
M 2 WTri 4.65–4.82* — 3.92–4.49 4.05–4.22 2.85–3.90 WTal 4.40–4.42 3.7 3.48–4.34 3.77–3.82 2.85–3.75
L 5.42–5.43 — 4.12–4.81 4.49 3.15–4.35
M 3 WTri 4.08–4.14 — 2.94–3.50 3.33 2.40–3.15 WTal 3.35–3.36 — 2.11–3.09 2.64 — L — — 3.75–4.30 — —
M 1 W — — 5.16–5.91 — —
L 5.37 — 3.88–4.34 — — M 2
W 6.65 — 5.68–6.49 — —
L 3.70 — 2.60–2.95 — — M 3
W 5.52 — 3.81–4.64 — —
* Estimate.
1 Measurements of P. pipiringosi from Gazin (1956) and P. acolytus from Rigby (1980: table 46). The holotype of P. lemuroides is AMNH 2421. Other specimens of P. lemuroides measured were: AMNH 15952, 15953, 15955, 15957, 15958, 15959, 16631, 16632, 16633, 16634, 16645 (lower dentitions) and AMNH 16636, 16644, 17045 (upper dentitions).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.