
Cyrtodactylus australotitiwangsaensis, Grismer, Lee, Wood, Perry L., Quah, Evan S. H., Anuar, Shahrul, Muin, Abdul, Sumontha, Montri, Ahmad, Norhayati & Bauer, Aaron M., 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.211847 |
|
DOI |
https://doi.org/10.5281/zenodo.6181175 |
|
persistent identifier |
https://treatment.plazi.org/id/593687E8-C120-825A-FF1E-F97CFB62FE97 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyrtodactylus australotitiwangsaensis |
| status |
sp. nov. |
Cyrtodactylus australotitiwangsaensis sp. nov.
Southern Titiwangsa Bent-toed Gecko Figs. 17 View FIGURE 17 , 18 View FIGURE 18
Cyrtodactylus pulchellus Lim et al. 2002:51 ; Grismer 2011:420 (in part).
Holotype.—Adult male ( ZRC 2.6979) collected on 27 August 2006 by L. Grismer. P. Wood, Jr., and T. Youmans on the back road up to Fraser’s Hill, Pahang, Peninsular Malaysia (03° 43.1699 N, 101° 45.4789 E) at 1059 meters above sea level.
Paratypes.— Paratypes ZRC 2.6980 and LSUHC 8088–90 (adult female and adult males, respectively) have the same collection data as the holotype. Paratype LSUHC 6637 (adult male) was collected on 18 August 2004 by P. Wood, Jr., and T. Youmans at Genting Highlands, Pahang, Peninsular Malaysia (03° 25.2388 N, 101° 47.3300 E) at 1346 meters above sea level.
Diagnosis.—Adult males reaching 120.1 mm SVL, adult females reaching 97.2 mm SVL; 9–12 supralabials, 9–13 infralabials; tubercles of dorsum small to moderately large with no intervening smaller tubercles; no tubercles on ventral surfaces of forelimbs, gular region, or in ventrolateral body fold; 37–45 paravertebral tubercles; 22–30 longitudinal rows of dorsal tubercles; 32–40 rows of ventral scales; 21–25 subdigital lamellae on fourth toe; 39–45 femoro-precloacal pores in males; dorsum not bearing a scattered pattern of white tubercles; three or four body bands in adults lacking lightened centers and light colored tubercles; band to interspace ratio 1.00–2.00; seven or eight dark, caudal bands on original tail; white caudal bands in adults immaculate; and posterior portion of tail in hatchlings and juveniles banded not white. These characters are scored across all species of the Cyrtodactylus pulchellus complex in Table 6 View TABLE 6 .
Description of holotype.—Adult male SVL 118.2 mm; head large, moderate in length (HL/SVL 0.28) and wide (HW/HL 0.73), somewhat flattened (HD/HL 0.37), distinct from neck, and triangular in dorsal profile; lores concave anteriorly, inflated posteriorly; frontal and prefrontal regions deeply concave; canthus rostralis sharply rounded anteriorly; snout elongate (ES/HL 0.40), rounded in dorsal profile, laterally constricted; eye large (ED/HL 0.22); ear opening elliptical, moderate in size (EL/HL 0.06), obliquely oriented; eye to ear distance greater than diameter of eye; rostral rectangular, divided dorsally by an inverted Y-shaped furrow, bordered posteriorly by left and right supranasals, and three small medial postrostrals (=internasals), bordered laterally by first supralabials; external nares bordered anteriorly by rostral, dorsally by a large anterior supranasal and small posterior supranasal, posteriorly by two smaller postnasals, ventrally by first supralabial; 11(R,L) rectangular supralabials extending to just beyond upturn of labial margin, tapering abruptly below midpoint of eye; first supralabial largest; 11(R) 10(L) infralabials tapering in size posteriorly; scales of rostrum and lores flat, larger than granular scales on top of head and occiput, those on posterior portion of canthus rostralis slightly larger; scales of occiput and top of head between eyes intermixed with small tubercles; large, boney frontal ridges bordering orbit confluent with boney, V-shaped, transverse, parietal ridge; dorsal superciliaries elongate, smooth, largest anteriorly; mental triangular, bordered laterally by first infralabials and posteriorly by left and right, rectangular postmentals which contact medially for 50% of their length; single row of enlarged, elongate sublabials extending posteriorly to seventh infralabial; small, granular to flat, gular scales grading posteriorly into larger, flat, smooth, imbricate, pectoral and ventral scales.
Body relatively short (AG/SVL 0.47) with well-defined, non-tuberculate, ventrolateral folds; dorsal scales small, granular, interspersed with larger, trihedral, regularly arranged, keeled tubercles, smaller intervening tubercles absent; tubercles extend from top of head from between eyes to caudal constriction and onto tail where they occur in transverse rows separated by six or seven small, flat scales; caudal tubercles largest dorsally, absent laterally and ventrally; caudal tubercles decrease in size posteriorly; tubercles on occiput and nape small, those on body largest; approximately 30 longitudinal rows of tubercles at midbody; 45 paravertebral tubercles; 38 flat imbricate ventral scales between ventrolateral body folds, ventral scales larger than dorsal scales; precloacal scales large, smooth; distinct precloacal groove.
Forelimbs moderate, relatively short (FL/SVL 0.18); scales on dorsal surfaces of forelimbs subimbricate, intermixed with larger tubercles; scales of ventral surface of forearm flat, subimbricate, lacking tubercles; palmar scales rounded; digits well-developed, inflected at basal, interphalangeal joints; subdigital lamellae rectangular proximal to joint inflection, only slightly expanded distal to inflection; digits more narrow distal to joints; claws well-developed, sheathed by a dorsal and ventral scale; hind limbs more robust than forelimbs, moderate in length (TBL/SVL 0.20), larger tubercles on dorsal surface of thigh separated by smaller granular scales, tubercles on dorsal surfaces of foreleg same size as those on thigh; ventral scales of thigh flat, smooth, imbricate, larger than dorsal granular scales; ventral, tibial scales flat, smooth, imbricate; single row of greatly enlarged, flat, rectangular, imbricate, pore-bearing femoral scales extend from knee to knee through precloacal region where they are continuous with enlarged, pore-bearing precloacal scales; 43 contiguous, pore-bearing femoro-precloacal scales forming an inverted T bearing a deep, precloacal groove in which eight pore-bearing scales are found (four on each side of groove); postfemoral scales immediately posterior to pore-bearing scale row smaller, forming an abrupt union with pore-bearing postfemoral scales on posteroventral margin of thigh; plantar scales low, slightly raised; digits well-developed, inflected at basal, interphalangeal joints; subdigital lamellae proximal to joint inflection rectangular, only slightly expanded distal to inflection; digits more narrow distal to joints; claws well-developed, sheathed by a dorsal and ventral scale; 24(R,L) subdigital lamellae on 4th toe.
Tail 146 mm in length, 9.8 mm in width at base, tapering to a point; dorsal scales of tail flat, squarish; tail segmented with six or seven transverse scale rows per segment; posterior margin of segments bordered by four larger tubercles dorsally in anterior one-third of tail, fewer posteriorly; subcaudal region bearing large, transverse scales; shallow dorsal and lateral caudal furrows extend entire length of tail; base of tail bearing hemipenial swellings; three small, postcloacal tubercles on hemipenial swellings; postcloacal scales smooth, flat, large, imbricate.
Coloration in life. Dorsal ground color of head, body, limbs, and tail dark beige; no white chevron on rostrum; wide, dark brown nuchal band extends from posterior margin of one eye to posterior margin of other eye, edged with prominent thin, cream-colored, lines; three wide, similarly colored dorsal bands between limb insertions with immaculate, white tubercles forming a dotted line edging the bands; first band terminates at shoulders, second band terminates just dorsal to ventrolateral fold midway between limb insertions, third band terminates in groin and on anterior margin of hind limb; body band/interspace ratio of 1.25; one additional dark brown band posterior to hind limbs; no band on posterior margin of thigh; seven wide, dark bands approximately twice the width of seven white bands extend onto tail; all bands encircle tail, no bands on tail or body have lighten centers or encompass yellowish to white tubercles; ventral surfaces of head, limbs, and tail dull white; gular scales stippled; abdomen immaculate, beige; weakly stippled ventral scales.
Variation. The dorsal color of the paratypes closely matches that of the holotype although there is some departure in pattern ( Fig. 18 View FIGURE 18 ). The second and third body bands of LSUHC 6637 are broken just left of the midline and are offset. The second body band in LSUHC 8090 and ZRC 2.6980 is broken laterally with a fragment occurring on the right and left side of the midline respectively. LSUHC 6637 and 8089 have four instead of three body bands LSUHC 8089 has a regenerated tail, and LSUHC 6637 has eight instead of six dark caudal bands. Meristic differences in the type series are presented in Table 13.
Additional specimens examined. Variation in meristic characters for the type series and the additional specimens examined is listed in Table 13. Juveniles have a much bolder and contrasted color pattern than adults ( Fig. 18 View FIGURE 18 ).
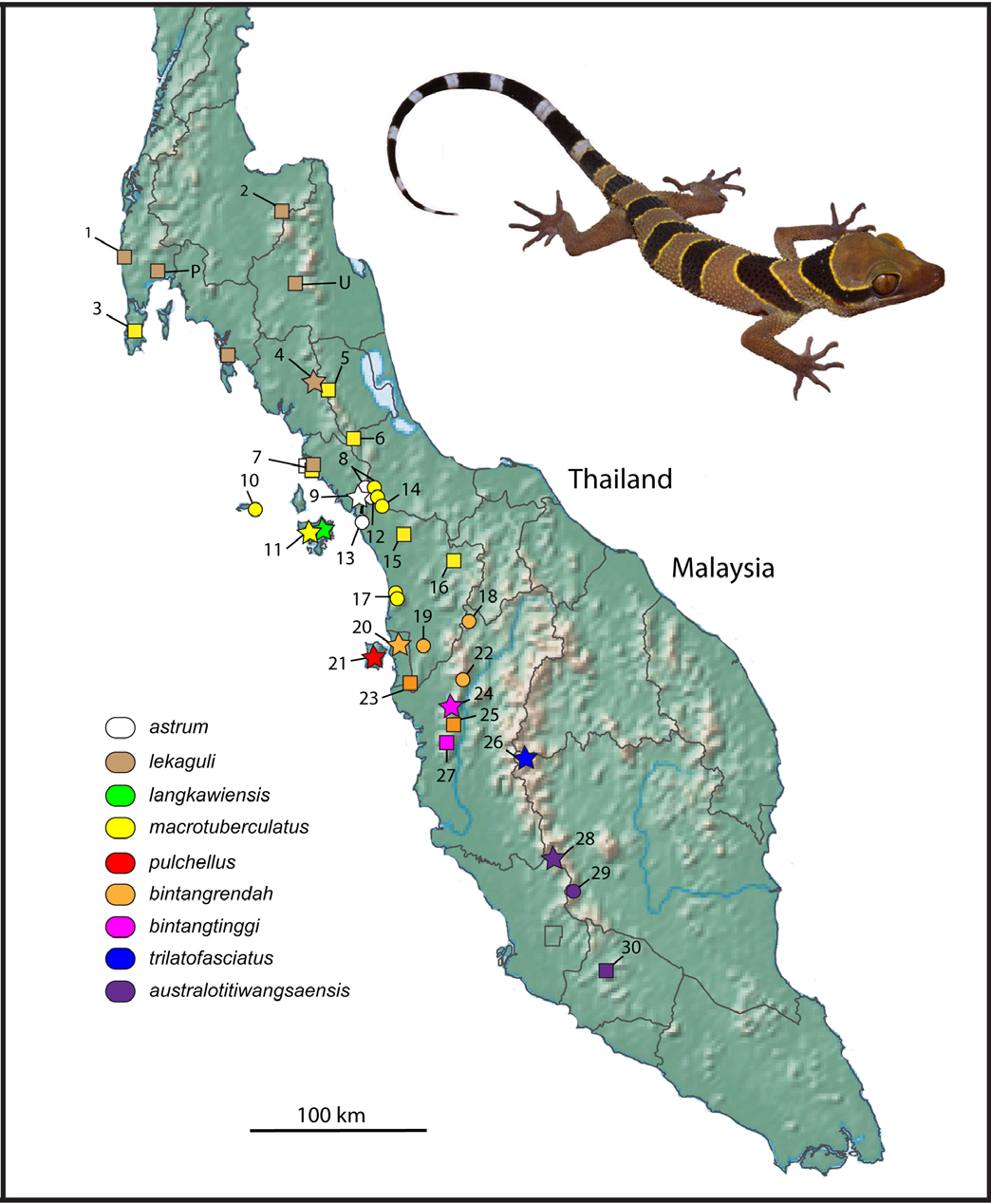
Distribution. Cyrtodactylus australotitiwangsaensis sp. nov. ranges along the upland areas of the southern portion of the Titiwangsa Mountains from at least Fraser’s Hill, Pahang in the north to Gunung Angsi, Negeri Sembilan in the south. Presumably it ranges farther to the north where it is likely to come in contact with its sister species C. trilatofasciatus sp. nov. and as far south perhaps as Gunung Ledang, Johor.
Natural history. Cyrtodactylus australotitiwangsaensis sp. nov. occurs in hill dipterocarp and lower montane forests ( Fig. 17 View FIGURE 17 ) from approximately 1000–1500 meters in elevation. It is a saxicolous lizard that emerges at night to forage among rocks or on nearby vegetation. At Fraser’s Hill and Genting Highlands, Pahang, lizards are quite common along rocky road cuts. This species does not occur in forested areas where rocks are absent. Juveniles from Fraser’s Hill have been collected in June ( Grismer 2011).
Etymology. The specific epithet adjective australotitiwangsaensis is derived from the combination of the Latin australis meaning southern and the name of the mountain range, Titiwangsa, in which this species is endemic and refers to its occurrence in the southern portion of this range.
Comparisons. Cyrtodactylus australotitiwangsaensis sp. nov. is separated from C. macrotuberculatus in lacking large tubercles on the dorsal surface of the head, body and limbs, on the underside of the forearms, in the gular region, and in the ventrolateral body fold, and in having 32–40 ventral scales vs. 17–28. It is separated from C. astrum sp. nov., C. langkawiensis sp. nov., C. lekaguli sp. nov., and C. pulchellus in having more than 38 femoro-precloacal pores (it overlaps with C. pulchellus by one pore at 39). It differs from C. trilatofasciatus sp. nov. with a body band/interspace ratio of 1.00–2.00 (three specimens [LSUHC 9107–08, ZRC 2.5396] out of 13 had a ratio of 2.00) vs. 2.00–2.75. Cyrtodactylus australotitiwangsaensis sp. nov. is further separated from C.
astrum sp. nov. in lacking a scattered dorsal pattern of white tubercles. Having hatchlings and juveniles that lack white tail tips separate it from C. astrum sp. nov., C. lekaguli sp. nov., and C. langkawiensis sp. nov. Having less than nine dark caudal bands differentiates C. australotitiwangsaensis sp. nov. from C. astrum sp. nov., C. langkawiensis sp. nov., and C. lekaguli sp. nov. It is further separated from C. astrum sp. nov., C. langkawiensis sp. nov., C. lekaguli sp. nov., and C. macrotuberculatus by having immaculate white caudal bands as adults as opposed to white bands that are infused with dark pigment. Its large size (maximum SVL 120.1 mm) separates it from C. astrum sp. nov., C. bintangtinggi sp. nov., C. langkawiensis sp. nov., and C. lekaguli sp. nov. in that these species do not reach over 111.1 mm SVL.
Cyrtodactylus macrotuberculatus Grismer & Norhayati 2008 Large Tubercled Bent-toed Gecko
Fig. 19 View FIGURE 19
Diagnosis.—Adult males reaching 118.6 mm SVL, adult females reaching 113.4 mm SVL; 9–12 supralabials, 7–11 infralabials; tubercles of dorsum very large with no intervening smaller tubercles; tubercles on ventral surfaces of forelimbs, in gular region, and in ventrolateral body fold; 31–44 paravertebral tubercles; 19–27 longitudinal rows of dorsal tubercles; 17–28 rows of ventral scales; 20–24 subdigital lamellae on fourth toe; 28–40 femoro-precloacal pores in males; dorsum not bearing a scattered pattern of white tubercles; four body bands in adults lacking lightened centers and light colored tubercles; band to interspace ratio 1.00–1.50; 7–10 dark, caudal bands on original tail; white caudal bands in adults infused with dark pigment; and posterior portion of tail in hatchlings and juveniles banded not white. These characters are scored across all species of the Cyrtodactylus pulchellus complex in Table 6 View TABLE 6 .
Remarks. Cyrtodactylus macrotuberculatus was sampled from the type locality on Langkawi Island and five additional peninsular localities including Perlis State Park and Bukit Chabang, Perlis; and Gunung Jerai (including Hutan Lipur Sungai Tupah), Kedah ( Fig. 3 View FIGURE 3 ). Although the Langkawi population is monophyletic (exclusive), its overall sequence divergence from the remaining populations is relatively modest (2.2–5.9%) and it is not morphologically diagnosable from them ( Tables 14–16 View TABLE 14 View TABLE 15 View TABLE 16 ). Although all four populations form a strongly supported monophyletic group, the relationships between them remain unresolved as indicated by the low support values from all three analyses ( Fig. 1). Therefore at this point, we refrain from considering them different species. Additionally, the relationships between the peninsular populations was somewhat inconsistent with their pattern of distribution given that a Gunung Jerai specimen (LSUHC 9693) grouped with samples from Bukit Chabang, precluding exclusivity (reciprocal monophyly) between these lineages as well as the polytomus relationships of the three major C. macrotuberculatus lineages ( Fig. 1). We consider the failure of haplotypes from a given locality to cluster together as evidence for potential gene flow between these localities. Lastly, these two groups are not morphologically diagnosable from one another ( Tables 14–16 View TABLE 14 View TABLE 15 View TABLE 16 ) and we know of geographically intervening populations that we were unable to sample and suspect that gene flow here is highly likely. As such, we leave the taxonomy of C. macrotuberculatus unchanged. With tissue samples and specimens from the Thai populations, additional taxonomic interpretations may be necessary even though at this point, the few samples of the Thai populations ( Tables 14–16 View TABLE 14 View TABLE 15 View TABLE 16 ) are not morphologically distinct from the Malaysian populations.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cyrtodactylus australotitiwangsaensis
| Grismer, Lee, Wood, Perry L., Quah, Evan S. H., Anuar, Shahrul, Muin, Abdul, Sumontha, Montri, Ahmad, Norhayati & Bauer, Aaron M. 2012 |
Cyrtodactylus pulchellus Lim et al. 2002 :51
| Grismer 2011: 420 |
| Lim 2002: 51 |