
Leucandra serrata Azevedo & Klautau, 2007
|
publication ID |
https://doi.org/10.11646/zootaxa.4363.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:7B5AC657-18B1-4A90-9268-FFAD49E1B9D0 |
|
DOI |
https://doi.org/10.5281/zenodo.6040372 |
|
persistent identifier |
https://treatment.plazi.org/id/5A058780-FFD7-A34D-13F7-7597F6D9FEB6 |
|
treatment provided by |
Plazi |
|
scientific name |
Leucandra serrata Azevedo & Klautau, 2007 |
| status |
|
Leucandra serrata Azevedo & Klautau, 2007 View in CoL
Synonymies. Leucandra serrata: Azevedo & Klautau 2007: 11 ; Muricy et al. 2008: 126; Lanna et al. 2009: 16; Ignacio et al. 2010: Table S1; Muricy et al. 2011: 29.
Type material. UFRJPOR 4878 ( holotype /alcohol), Angra dos Reis ( Bonfim Island ) ; collected by M. Klautau ( 5 April 2003), 2 m depth. UFRJPOR 4879, UFRJPOR 4881 ( paratypes /alcohol), Angra dos Reis ( Botinas Islands ) ; collected by M. Klautau ( 5 April 2003), 2 m depth.
Type locality. Bonfim Island , Angra dos Reis, Rio de Janeiro, Brazil.
Analysed material. UFBA 4525 - POR and UFBA 4658 - POR [ Marina of the Nautical Tourist Terminal of Bahia (1258’20.8’’S, 3830’54.6’’W), Salvador , Bahia State, Brazil; collected by C. Chagas; 20/II/2016; 1 m depth], UFRJPOR 4887 ( Angra dos Reis, Botinas Islands; collected by M. Klautau & E. Lanna, 17 April 2004, 3 m depth), and holotype .
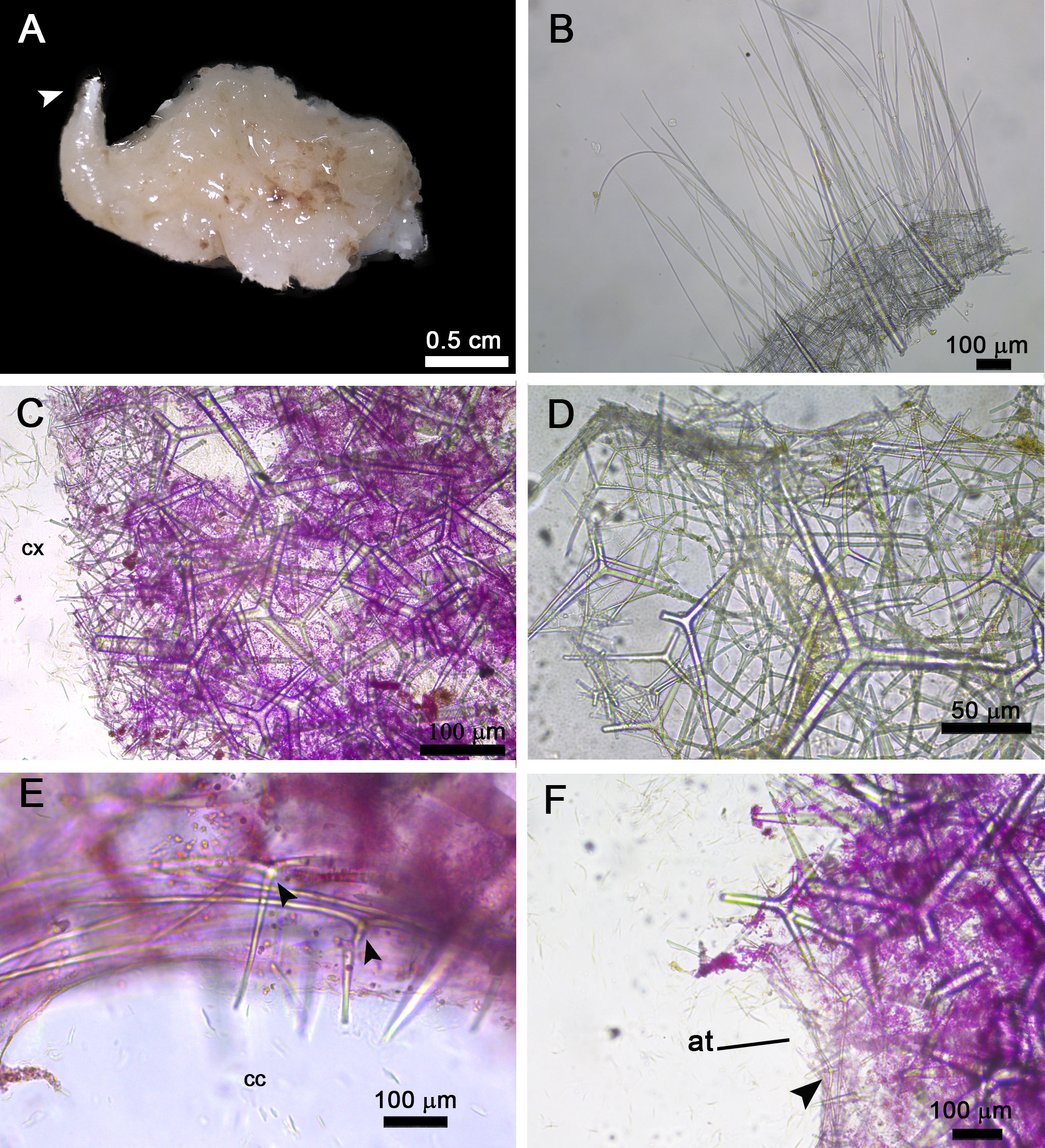
Description: Colour is beige alive and after fixation. The largest specimen from Salvador measures 1.2 x 2.1 cm (height x width) and has an irregular shape ( Figure 2A View FIGURE2 ). The osculum is apical and has a membrane filled with spicules, to which several trichoxeas are inserted forming a fringe ( Figure 2B View FIGURE2 ). The surface of the specimens is undulated and hispid, with some diactines perforating the cortex. The body wall is 0.5 cm thick. The aquiferous system is leuconoid, with ovoid choanocyte chambers. The atrial cavity fills about half of the specimens.
The skeleton of the cortex is formed mainly by triactines of different shapes and sizes and by diactines that penetrate the cortex and extend until the choanosomal region ( Figures 2C, D View FIGURE2 ). Microdiactines are also present at the sponge surface ( Figure 3B View FIGURE 3 ). The choanosomal skeleton is formed by large triactines arranged in a disorganised way ( Figure 2C View FIGURE2 ). Additionally, we found tetractines around the canals, their apical actines facing the lumen ( Figure 2E View FIGURE2 ). These spicules are smaller than the choanosomal triactines. The atrial skeleton is formed by tetractines with the apical actine projected to the atrium ( Figure 2F View FIGURE2 ).
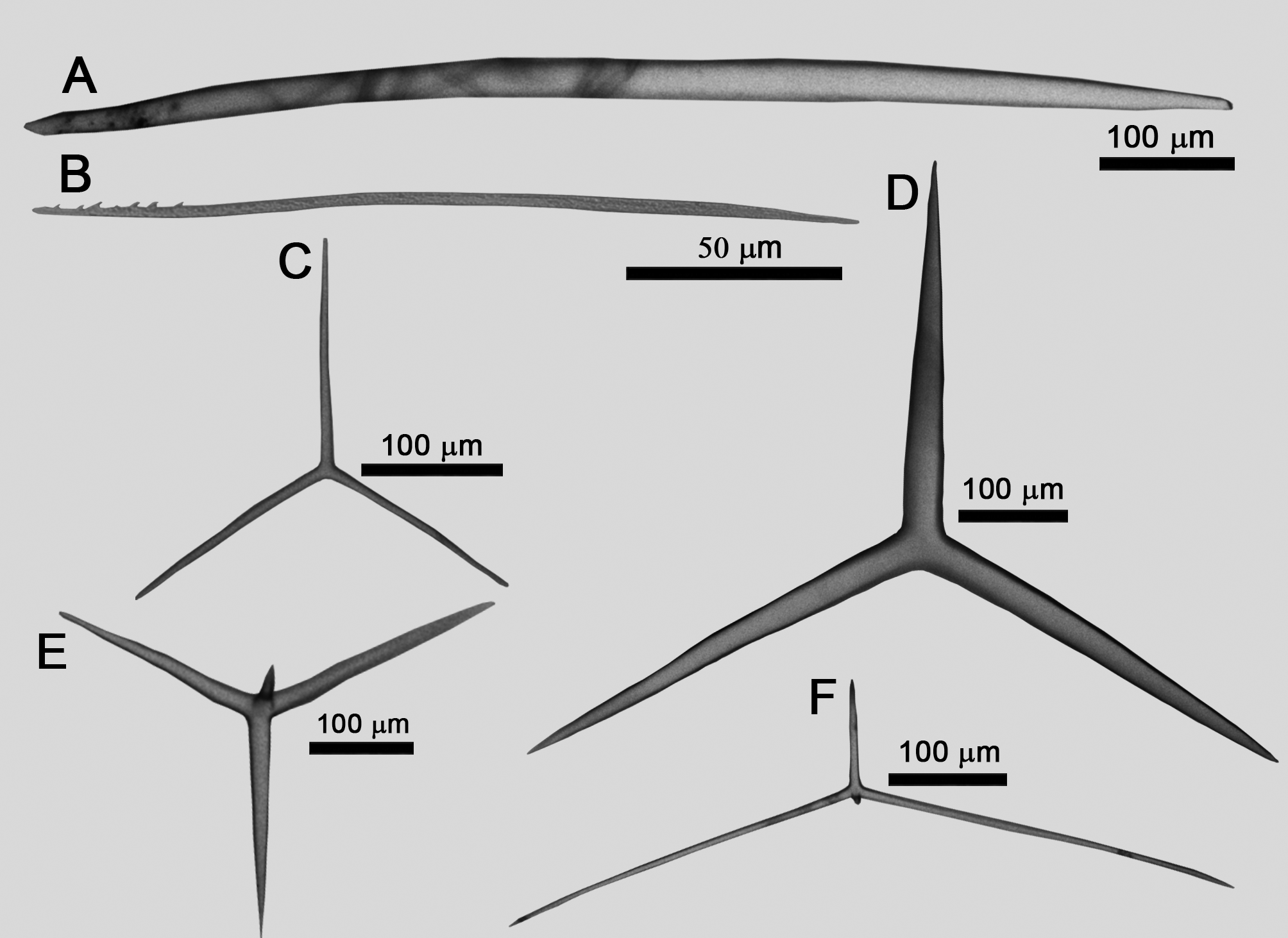
Spicules ( Table 1; measurements obtained from two specimens):
Trichoxeas: Very thin and long. Most of them are broken.
Diactines: Fusiform and sharply pointed or with one of the tips blunt [478.8– 1023.0 ( ± 287.5)–1769.0/15.9– 42.8 ( ± 12.4)–61.5 µm; Figure 3A View FIGURE 3 ]
Microdiactines: Thin, slightly curved and jagged at one of the tips [71.3– 107.6 (24.0)–170.2/2.1– 4.3 (1.3)–7.0 µm; Figure 3B View FIGURE 3 )]
Cortical triactines: Cylindrical with blunt tips. The paired actines are commonly curved. They have the same size or are larger than the unpaired one [paired: 12.6– 199.1 (48.5)–285.7/3.7– 7.3 (2.1)–16.1 µm; unpaired: 42.9– 131.1 (55.9)–288.7/3.9– 8.0 (2.0)–14.3 µm; Figure 3C View FIGURE 3 ].
Choanosomal triactines: Conical and sharp pointed. The size is variable, but they are the largest spicules present in the skeleton. The paired actines are slightly larger than the unpaired one and they are also sometimes slightly curved. In some cases, all actines have different sizes [paired: 177.3– 490.5 (198.7)–948.9/14.5– 37.4 (13.8)–65.5 µm; unpaired: 128.0– 352.1 (133.4)–667.0/15.1– 38.3 (15.1)–67.7 µm; Figure 3D View FIGURE 3 ).
Choanosomal tetractines (canals): Conical with sharp tips. Some spicules have a slight undulation close to the end of one of the paired actines. The unpaired actine is smaller than the paired ones. The apical actine is short [paired: 107.5– 210.4 (46.2)–310.0/4.8– 10.9 (3.0)–18.2 µm; unpaired: 84.1– 184.2 (42.4)–279.2/7.4– 11.6 (2.7)– 21.2 µm; apical: 17.0– 42.4 (14.5)–86.2/4.5– 8.2 (2.1)–15.9 µm; Figure 3E View FIGURE 3 ].
Atrial tetractines: Cylindrical and with sharp tips. Paired actines are slightly curved and long, being larger than the other actines. The apical is short but sometimes it is larger than the unpaired actine (paired: 122.3– 243.5 (32.9)–303.8/5.0– 8.6 (2.2)–13.6 µm; unpaired: 50.4– 100.6 (24.6)–195.7/4.3– 9.3 (2.2)–14.5 µm; apical: 9.5– 58.4 (22.5)–110.7/4.0– 8.0 (2.2)–14.3µm; Figure 3F View FIGURE 3 ].
Ecology. The artificial substrates where the specimens were sampled remained immersed for 2 months, so this is the maximum age of the sponges. Tunicates, bryozoans and demosponges, as well as macroalgae, were also among the colonisers. Considering the bathymetric distribution, it seems that L. serrata is from very shallow water, as, to our knowledge, it occurs from 1 to 3 m in depth ( Azevedo & Klautau 2007; Lanna et al. 2009; current work).
Remarks. Leucandra serrata was originally described at the coast of Angra dos Reis, Rio de Janeiro state ( Azevedo & Klautau 2007) and was subsequently recorded 2.900 km away, in the Potiguar Basin, Northeastern Brazil ( Muricy et al. 2008; Lanna et al. 2009). The presence of this species in the coast of Bahia fills a large gap in the distribution of L. serrata along the Brazilian coast, as Bahia is between those localities.
The specimens described here have jagged microdiactines, long diactines in the cortex and large triactines at the choanosomal region, which are characteristics mentioned by Azevedo & Klautau (2007) and Lanna et al. (2009) for L. serrata . The only difference is the size of the choanosomal triactines: in the specimens from Bahia they are larger (318.9– 625.0 (185.5)–948.9/49.0 (8.7) µm) than those from the Potiguar Basin (156.0– 294.0 (63.9)–426/26.8 (6.0) µm) and from Rio de Janeiro (234.0– 420.0 (69.0)–523.0/30.0 (6.0) µm).
| POR |
Universit� degli Studi di Napoli Federico II |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |