
Correlophus
|
publication ID |
https://doi.org/ 10.5281/zenodo.211734 |
|
DOI |
https://doi.org/10.5281/zenodo.6166578 |
|
persistent identifier |
https://treatment.plazi.org/id/5C4F87E1-FFA8-FFB1-088F-F931D293B1A9 |
|
treatment provided by |
Plazi |
|
scientific name |
Correlophus |
| status |
|
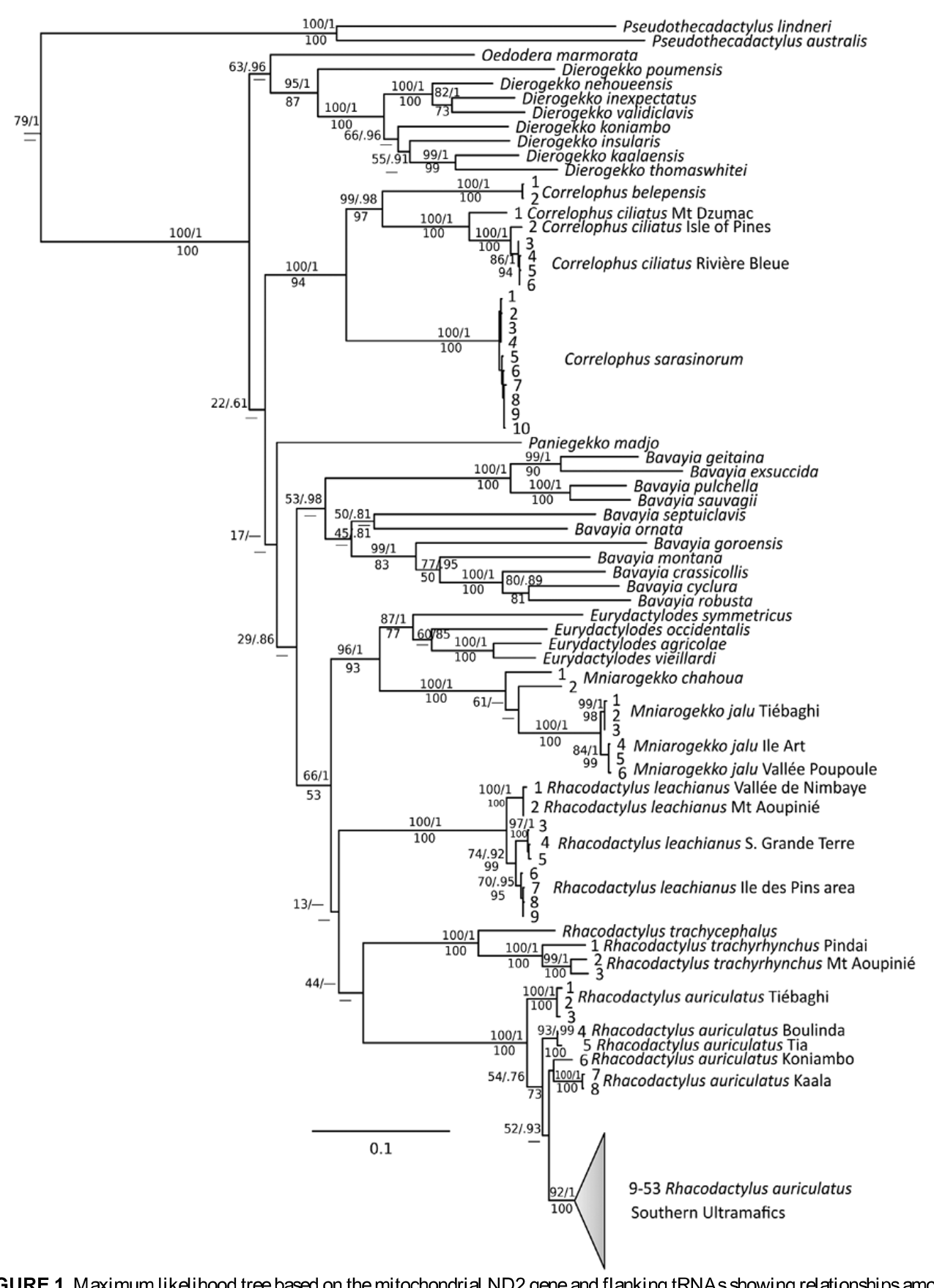
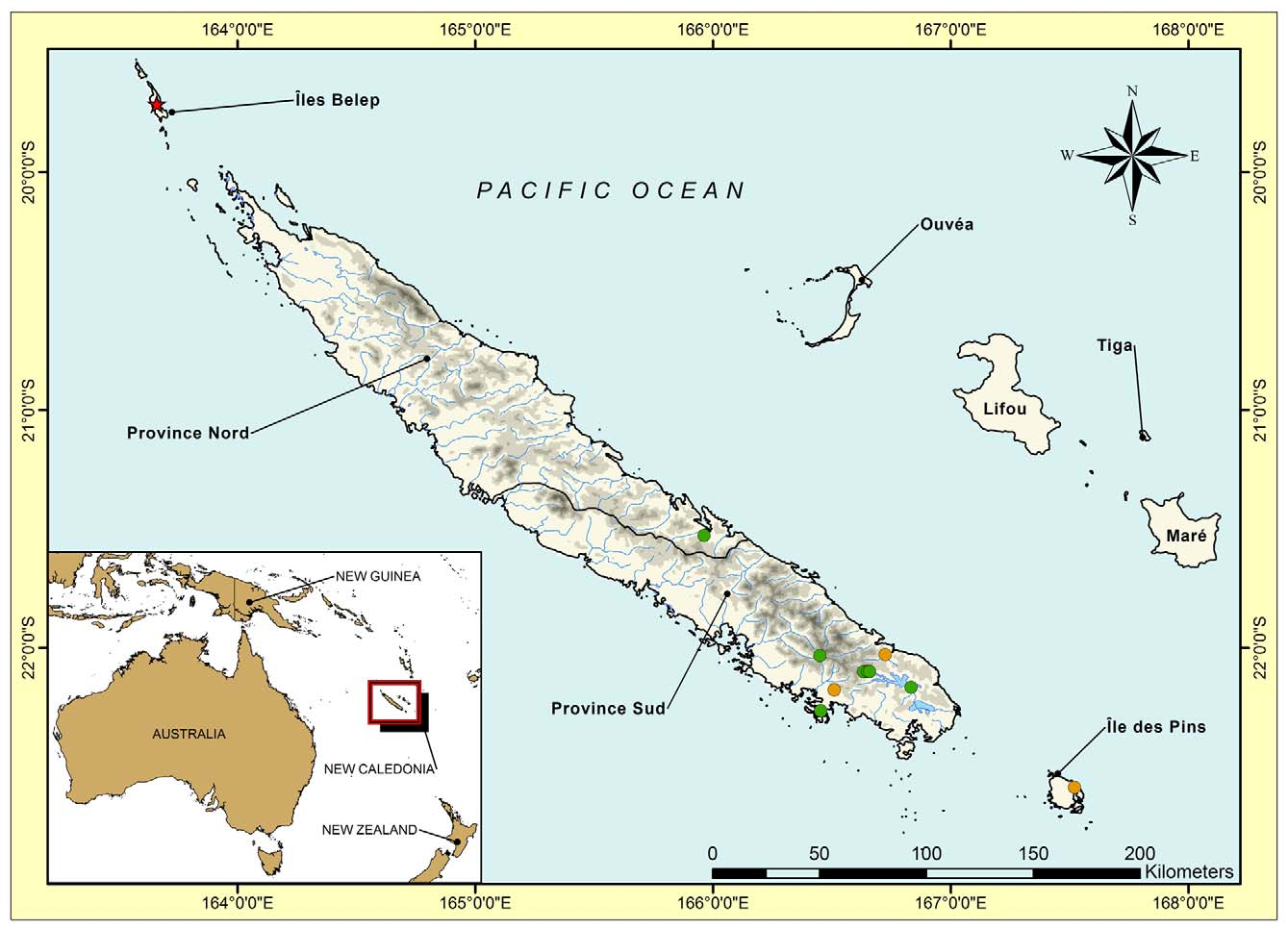
Correlophus sarasinorum exhibits almost no intraspecific variation in the genetic markers we studied, even in the rapidly evolving mitochondrial genes ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ). This is perhaps not surprising given the very restricted distribution of this species, which is limited to the southern ultramafic block of the Grande Terre (Bauer 1990; Bauer & Henle 1994; Bauer & Sadlier 2000; Fig. 9 View FIGURE 9 ). Within this limited variation specimens from Fôrét Nord in the far south of the Plaine des Lacs differ only minimally from those further north at Mt. Koghis and Bois du Sud. Variation in color pattern has been previously mentioned ( Böhme & Henkel 1985; Henkel 1987, 1988; Bauer 1990; Myers 1997) but this appears to have no obvious phylogenetic basis. Good et al. (1997) found four fixed allozyme differences between single individuals of C. sarasinorum from Touaourou and Rivière Bleue, leading Bauer and Sadlier (2001) to hypothesize that two species might be represented. The information from the DNA sequence data presented here clearly contradicts this assumption and highlights the pitfalls of limited sampling. Several “morphs” are recognized by hobbyists (de Vosjoli et al. 2003), but these are also of no phylogenetic significance.
Correlophus ciliatus —The greatest intraspecific genetic divergence within any giant gecko was seen within C. ciliatus ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ). This species was described in 1866 ( Fig. 10 View FIGURE 10 ) and was apparently not uncommon in that era ( Bavay 1869). It was then “lost” to science for over 100 years and considered likely to be extinct ( Bauer & Sadlier 1993) until rediscovered on the Île des Pins in the 1990s ( Storelli 1994; Seipp & Klemmer 1994; Kullmann 1995). It was subsequently found on several smaller satellite islands around Île des Pins (de Vosjoli 1995) and in the southern Grande Terre ( Girard & Heuclin 1998; Bauer & Sadlier 2000, 2001; Fig. 11 View FIGURE 11 ). Since then it has become one of the most popular of all lizard pets and is bred in at least the tens of thousands around the world ( Baldwin & Repashy 1998; Both 1999; Bach 2006). All or most of these animals appear to originate from the Île des Pins, rather than Grande Terre. We found very little divergence between Île des Pins specimens and those from Rivière Bleue on the mainland, but quite deep divergences, comparable to the deepest within R. auriculatus , between these and a single specimen from Mt. Dzumac, only about 20 km distant from Rivière Bleue. However, specimens from the recently discovered population from the Îles Belep ( Whitaker et al. 2004; Wirth & Peukert 2009) were as divergent as the most deeply-divergent splits between species in the Bavayia sauvagii clade.
Further, there are morphological differences between the southern and northern populations concordant with the genetic differences retrieved, lending futher support to the recognition of the Belep Island populations as an independent evolutionary lineage, and on these criteria we here recognize this northern “ ciliatus ” as a new species:



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.