
Prionospio anneae, Radashevsky, Vasily I., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4019.1.22 |
|
publication LSID |
lsid:zoobank.org:pub:88F2DB05-58C4-4726-89D5-99302FABB908 |
|
DOI |
https://doi.org/10.5281/zenodo.4658152 |
|
persistent identifier |
https://treatment.plazi.org/id/5E51D737-FFD3-FFB0-FF4A-A4B71B36FD8B |
|
treatment provided by |
Plazi |
|
scientific name |
Prionospio anneae |
| status |
sp. nov. |
Prionospio anneae View in CoL n. sp.
( Figs 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Type material. Queensland: Holotype: AM W.45273, MI QLD 2410. Paratypes: AM W.45271, MI QLD 2330 (1); AM W.45272, MI QLD 2363 (1); AM W.47870, MI QLD 2373 (1); AM W.47871, MI QLD 2410 (2); AM W.45274, MI QLD 2431 (3); MIMB 28109, MI QLD 2431 (1).
Adult morphology. Up to 14 mm long, 0.3 mm wide for about 90 chaetigers; holotype 12 mm long for 53 chaetigers. Pigmentation on body absent; some yellow indistinct spots present on branchiae in life. Prostomium narrow and rounded anteriorly, extending posteriorly to middle of chaetiger 2 as a prominent caruncle. Occipital 1. The identification key is based on characters appearing in completely developed adults. For example, median eyes enlarge in ontogenesis apparently in association with sexual maturation of an individual, caruncle elongates with age up to a certain speciesspecific limit, pinnae on branchiae (especially on posterior pairs) may appear in a while after development of the branchiae, thus smooth, apinnate branchiae are present in small and median-size individuals on chaetigers where pinnate branchiae are present in completely developed adults. Identification of those small and median-size individuals is therefore problematic, unless ontogenetic development and transformation of characters is taken into account.
2. Branchiae pinnate only on chaetiger 2 in small individuals, on chaetigers 2 and 3 in median-size individuals.
3. Branchiae pinnate only on chaetiger 2 in small individuals, on chaetigers 2 and 4 in median-size individuals.
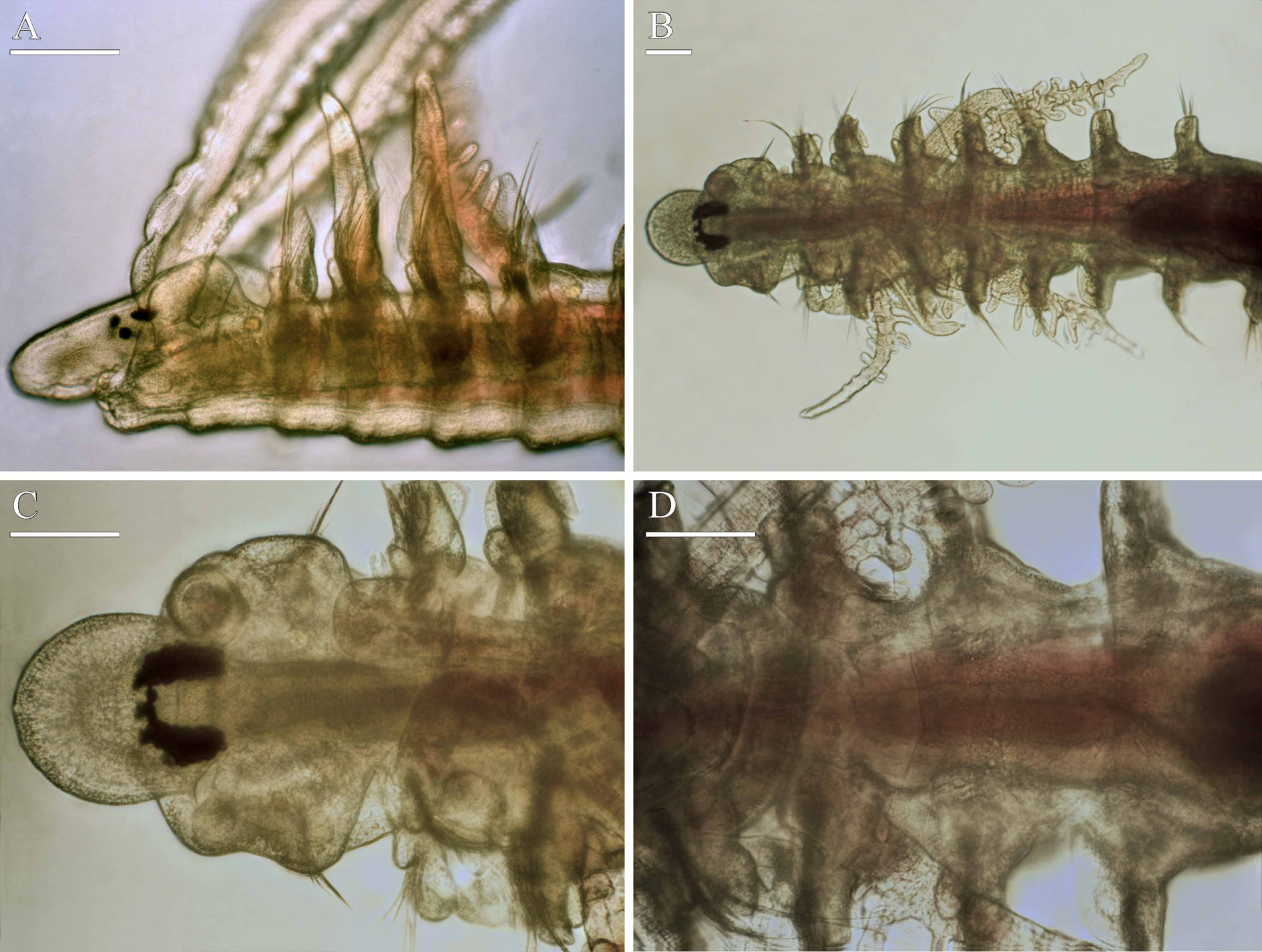
antenna absent. Two pairs of red eyes arranged trapezoidally, comprising one pair of median eyes and one pair of lateral eyes situated anteriorly and set wider apart; median eyes small to moderately large in individuals with 40–60 chaetigers, very large in bigger individuals ( Fig. 8 View FIGURE 8 A–C). Nuchal organs U-shaped ciliary bands on each side of caruncle. Posterior dorsal part of peristomium fused to notopodial lamellae of chaetiger 1 forming prominent rounded ear-shaped structures. Palps as long as 10–15 chaetigers, with frontal longitudinal groove lined with fine cilia, short transverse bands of cilia regularly arranged on inner surface, and narrow longitudinal band of cilia running on outer fronto-lateral side along frontal groove; cilia of transverse bands beating towards distal end of palp along palp axis, while cilia of longitudinal band beating towards frontal groove perpendicular to palp axis.
Chaetiger 1 in individuals with less than about 50 chaetigers having only few capillaries in neuropodia; bigger individual with few short capillaries and postchaetal lamellae in both rami; notopodial lamellae large, flat, fused to posterior dorsal part of peristomium forming prominent ear-shaped structures ( Figs 8 View FIGURE 8 A, 9A). Notopodial postchaetal lamellae largest on branchial chaetigers. Lower part of neuropodial postchaetal lamellae of chaetiger 2 elongated and rounded ventrally ( Fig. 9 View FIGURE 9 B). Neuropodial postchaetal lamellae trapezoidal on chaetiger 3, smaller and rounded on succeeding chaetigers ( Fig. 9 View FIGURE 9 C–G).
Moderate dorsal crest present on chaetiger 6 ( Fig. 9 View FIGURE 9 F); very low crests or folds present from chaetiger 7 ( Fig. 9 View FIGURE 9 G) to chaetigers 15–21 (dorsal crest on chaetiger 6 and folds on chaetigers 7–20 in holotype). Lateral pouches and ventral flaps absent.
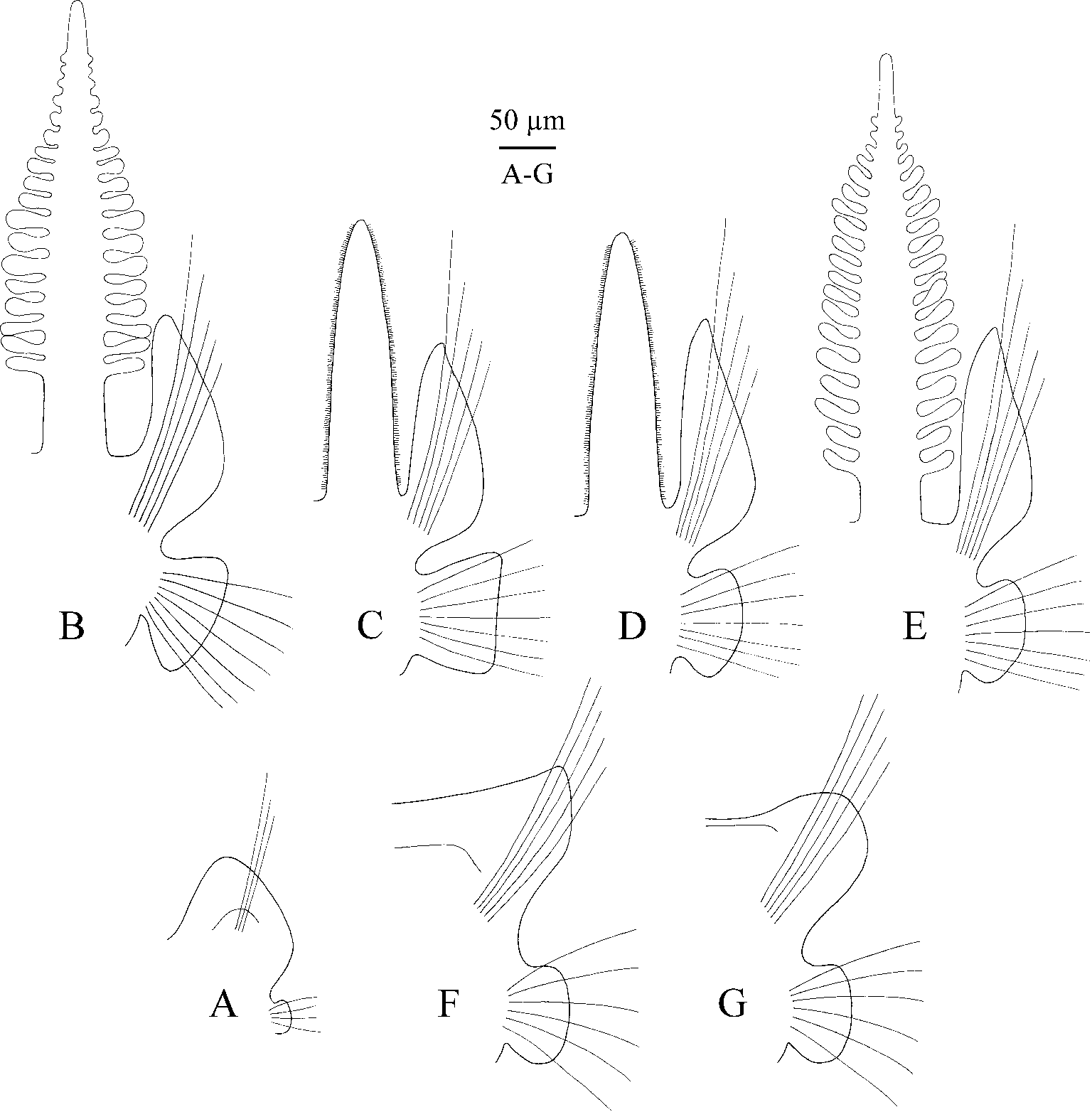
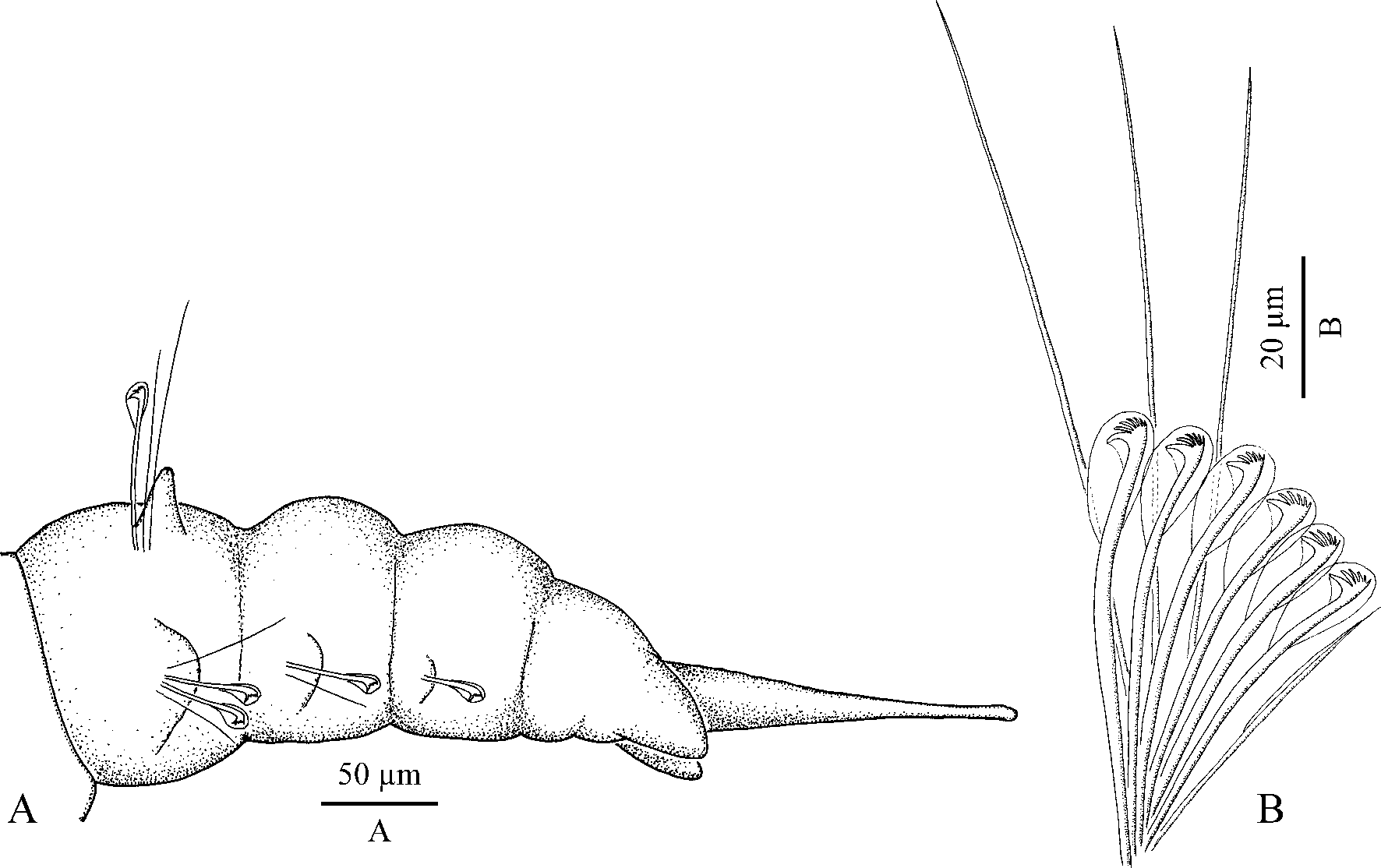
Hooks in notopodia from chaetigers 28–39 (39 in holotype), up to five in a series among capillaries. Hooks in neuropodia from chaetigers 8–9 (8 in holotype), up to eight in a series, accompanied by 1–2 inferior capillaries and 1–4 alternating capillaries throughout ( Fig. 10 View FIGURE 10 B). Inferior capillaries as long as hooks, alternating capillaries about two times as long as hooks; both kinds of capillaries in anterior chaetigers slender, with narrow limbation, in posterior chaetigers hair-like, alimbate. Hooks multidentate, with 4–6 pairs of thin, long upper teeth arranged in two vertical rows above main fang, with inner and outer hoods; shaft slightly bent. Sabre chaetae in neuropodia absent.
Four pairs of branchiae on chaetigers 2–5 ( Figs 8 View FIGURE 8 A, B, 9B–E); those on chaetigers 2 and 5 cylindrical, similar in length to each other, up to two times as long as notopodial lamellae, with numerous digitiform pinnae arranged along lateral and posterior sides along their length. Branchiae on chaetigers 3 and 4 up to 1.5 times as long as notopodial lamellae, apinnate, robust, flattened, with surface oriented perpendicular to body axis. An anterior fragment of a small individual about 0.15 mm wide with 2–3 pairs of small buds of pinnae on each of branchiae on chaetiger 2, and 7–8 pairs of well developed pinnae on each of branchiae on chaetiger 5. Longitudinal bands of cilia running on inner and outer edges on each branchia; ciliation heavier on branchiae on chaetigers 3 and 4. Afferent and efferent blood vessels interconnected by numerous radial capillaries forming loops inside individual pinnae.
Short nototrochs present on chaetigers 3 and 4, each composed by one transverse row of cilia. Dorso-lateral longitudinal ciliation present on chaetigers 3–5 as short bands of cilia extending between successive notopodia.
Pygidium with thin, long middorsal cirrus and one pair of short and thick ventral cirri (3–4 times as short as middorsal cirrus) ( Fig. 10 View FIGURE 10 A).
Oesophagus extending through 6–9 anterior chaetigers. Ventral buccal bulb below oesophagus extending to end of chaetiger 1. Gizzard-like structure in digestive tract absent.
Main dorsal blood vessel transformed into gut sinus in anterior part of midgut. Soft heart body up to 13 µm in diameter extending inside main dorsal vessel from level of chaetigers 3–4 to chaetigers 10–13. Blood red, without globules or other elements.
Nephridia in chaetigers 4–6, greenish in life.
Reproduction. Prionospio anneae n. sp. is gonochoristic. Four females have oocytes from chaetiger 10 to chaetigers 45–48, and two males have spermatozoa from chaetiger 11 to chaetigers 50–60. Oogenesis is intraovarian. Vitellogenic oocytes develop in ovaries attached to segmental blood vessels. Intraovarian oocytes were up to 75 µm in diameter, with germinal vesicle about 30 µm and single nucleolus 13 µm in diameter. Spermatogonia proliferate in testes; spermatogenesis occurs in the coelomic cavity. Spermatids are joined in tetrads. Spermatozoa are ect-aquasperm with small acrosome, spherical nucleus 2–3 µm in diameter, spherical mitochondria probably four in number, and a long flagellum.
Remarks. Prionospio with branchiae from segment 2, some apinnate and some with digitiform pinnules, have been treated as Prionospio Malmgren sensu stricto after Foster (1971) and Maciolek (1985). Those with four pairs of branchiae, pinnate on chaetigers 2 and 5, and apinnate on chaetigers 3 and 4, have conventionally been combined into the P. steenstrupi group of species named after P. steenstrupi described from northern Iceland by Malmgren (1867). Twelve species of this group occurring worldwide were summarized by Maciolek (1985: table 3, species 1–12). Imajima (1990a, b) described six new species from Japan. Hylleberg & Nateewathana (1991) recognized Prionospio malayensis Caullery, 1914 , originally described as a subspecies, as a valid species, and also described eight new species, members of the P. steenstrupi group, from the Andaman Sea. Sigvaldadóttir & Mackie (1993) recognized P. fallax Söderström, 1920 as a distinct species. Blake (1996) described two new species, P. jubata and P. marsupialia , from California. Zhou & Li (2009) described a new species, P. pacifica , from the South- China Sea, and Delgado-Blas (2015) described four new species from the Grand Caribbean Region.
In Australia, Blake & Kudenov (1978) reported P. steenstrupi from New South Wales and Victoria, and described two new species, P. australiensis and P. queenslandica , from Queensland. Hutchings & Rainer (1979) described a new species, P. multicristata , from New South Wales, and Wilson (1990) described three new species, members of the group, P. coorilla , P. kulin , and P. nirripa , from south-eastern Australia. Three new species, P. anneae n. sp., P. lylei n. sp. and P. cooki n. sp., with the same arrangement of branchiae as in P. steenstrupi are described in the present paper, thus making total number of species in the group 41. No identification key for the whole group is provided.
Prionospio anneae View in CoL n. sp. belongs to the P. steenstrupi View in CoL group and is characterized by having large median eyes, dorsal crest on chaetiger 6, hooks in neuropodia from chaetigers 8–9 and in notopodia from chaetigers 28–39, and inferior capillaries present in hook-bearing neuropodia instead of sabre chaetae. A low dorsal crest on chaetiger 6 is also present in only one other member of the P. steenstrupi View in CoL group, P. marsupialia Blake, 1996 View in CoL which differs from most of those species by having inter-neuropodial pouches. Inferior capillaries but not sabre chaetae are also present in neuropodia of P. perkinsi Maciolek, 1985 View in CoL and P. cerastae View in CoL n. sp. described below. These two species are, however, not members of the P. steenstrupi View in CoL group and have an arrangement of pinnate branchiae different from P. anneae View in CoL n. sp.
Etymology. The species is named in honour of Dr Anne Hoggett, a co-director (with her husband Dr Lyle Vail) of the Lizard Island Research Station, an avid enthusiast of marine biology and coral reef protection.
Habitat. Adult P. anneae n. sp. were found in coral sand and rubble at 5–16 m depth.
Distribution. Australia, Queensland, Great Barrier Reef.
| MIMB |
Museum of the Institute of Marine Biology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Prionospio anneae
| Radashevsky, Vasily I. 2015 |
P. marsupialia
| Blake 1996 |
P. perkinsi
| Maciolek 1985 |