
Srilankametrus serratus (Pocock, 1900), 2020
|
publication ID |
https://doi.org/10.1206/0003-0090.442.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/5E6CB374-FE5C-6CB9-FD73-62ECFEE9DB81 |
|
treatment provided by |
Felipe (2021-08-30 19:46:52, last updated 2024-07-19 09:14:50) |
|
scientific name |
Srilankametrus serratus (Pocock, 1900) |
| status |
comb. nov. |
Srilankametrus serratus (Pocock, 1900) View in CoL ,
comb. nov.
Figures 10 View FIGURE 10 , 28C, E, F View FIGURE 28 , 42C, E, F View FIGURE 42 , 57I–L View FIGURE 57 , 76F View FIGURE 76 , 77F View FIGURE 77 , 78F View FIGURE 78 , 198 View FIGURE 198 , 244 View FIGURE 244 , 247–251 View FIGURE 247 View FIGURE 248 View FIGURE 249 View FIGURE 250 View FIGURE 251 , table 1
Scorpio caesar: Pocock, 1894a: 73 (misidentification).
Heterometrus caesar: Kraepelin, 1899: 109 , 114 (part); Roewer, 1943: 229 (part); Takashima, 1945: 92 (part).
Palamnaeus caesar: Pocock, 1900a: 86 , 97 (part).
Palamnaeus serratus Pocock, 1900a: 86 , 97; Kovařík, 2004: 17, 19, 20, fig. 16.
Heterometrus serratus: Takashima, 1945: 94 ; Kovařík et at., 2016: 1, 5, 10, 103–108, 110, 111, 115, 117, 125, 126, figs. 15, 508–534, 571, 574, 593–595; 2019a: 12, fig. 53.
Heterometrus (Srilankametrus) indus: Couzijn, 1981: 82 , 94, 121, 123 (misidentification, part); Kovařík, 1998: 137, 138 (part); Fet, 2000: 446, 447 (part).
Heterometrus (Srilankametrus) indus indus: Couzijn, 1981: 39 , 82, 121–124, table 7 (misidentification, part); Fet, 2000: 447 (part).
Heterometrus (Srilankametrus) serratus: Tikader and Bastawade, 1983: 545 , 555–561, figs. 1483–1496; Fet, 2000: 448.
Heterometrus indus: Kovařík, 2004: 17 View Cited Treatment , 19, 20, 21, 32, 51, table 2 (misidentification, part), fig. 16; 2009: 35, 38, 48, table 1 (misidentification, part).
Heterometrus (Srilankametrus) servatus : Majum- der and Dey, 2005: 14 (misidentification).
TYPE MATERIAL: SRI LANKA: Holotype ♂ (BMNH 1888.55), Ceylon, Dr. Ondaatje [examined].
DIAGNOSIS: Srilankametrus serratus may be separated from other species of Srilankametrus as follows. The carapace is slightly dorsoventrally compressed, the lateral surfaces sloping moderately (fig. 28C, E, F) in S. serratus , but markedly dorsoventrally compressed, the lateral surfaces sloping gently, in S. indus and S. pococki , and vaulted, the lateral surfaces sloping steeply, in S. caesar , S. couzijni , S. gravimanus , and S. yaleensis . The interocular and posterior sutures are present, the posterior sutures extending past the median ocular tubercle and connected anteriorly to the posterior bifurcations of the interocular suture, in S. serratus , whereas the sutures are absent in S. indus and S. pococki . The carapace interocular surface is entirely smooth (fig. 28C, E, F) in S. serratus whereas the frontal lobes and medial region of the interocular surface are granular with smooth areas in S. caesar . The carapace anterolateral and mediolateral surfaces are granular in S. serratus but smooth or nearly so in S. indus . The carapace posterolateral surfaces of the female are granular in S. serratus but smooth or nearly so in S. gravimanus and S. yaleensis . The pedipalp patella dorsomedian carina of the female is absent or obsolete in S. serratus but entirely to predominantly costate in S. caesar . The patella retrodorsal carina of the female is as strongly developed as or more strongly developed than the retromedian carinae in S. serratus , but absent or obsolete in S. caesar , S. indus , and S. pococki . The retromedian carinae of the female are granular in S. serratus but absent or obsolete in S. gravimanus and S. yaleensis . The pedipalp chela of the adult male (figs. 248, 250) is sparsely setose in S. serratus but moderately to densely setose in S. indus and S. pococki . The chela manus dorsal surface (between the dorsomedian and digital carinae) is curved and slightly to markedly convex in S. serratus but flat in S. pococki . The proximal margin (lobe) of the dorsal surface is moderately curved and aligned with the proximal margin of the condyle (articulation with patella) (figs. 248, 250, 251) in S. serratus but markedly curved and proximal to the proximal margin of the condyle in S. couzijni . The dorsomedian carina is obsolete on the chela fixed finger and manus in S. serratus , pronounced and costate on the fixed finger and distally on the manus in S. caesar , S. couzijni , and S. gravimanus , and pronounced and costate on the fixed finger and distal threequarters of the manus in S. yaleensis . The maximum distance between the dorsomedian and dorsal secondary carinae (DMC–DSC) of the chela manus is greater than the maximum distance between the dorsal secondary and digital carinae (DSC–DC) in the male (figs. 248, 250) of S. serratus but similar to the DSC–DC in the male of S. gravimanus . The dorsal secondary and subdigital carinae of the male (figs. 248, 250) are entirely to predominantly granular in S. serratus but entirely to predominantly costate in S. gravimanus and S. yaleensis , and absent or obsolete in S. indus and S. pococki . The digital carina is entirely to predominantly granular (figs. 248, 250, 251) in S. serratus , entirely to predominantly costate in S. couzijni , S. gravimanus , and S. yaleensis , and absent or obsolete in S. indus and S. pococki . The retromedian carina of the male is entirely to predominantly granular in S. serratus , entirely to predominantly costate in S. couzijni , S. gravimanus , and S. yaleensis , and absent or obsolete in S. indus and S. pococki . The manus ventral surface is flat, with the axes of the retroventral and ventromedian carinae in approximately the same plane in S. serratus , but angled, with the axis of the retroventral carina ventral to the axis of the ventromedian carina, in S. indus and S. pococki . Macroseta st on the retroventral surfaces of the basitarsi of legs I and II is spiniform (fig. 57I–L) in S. serratus but setiform in all other species except S. caesar and S. gravimanus . The lateral surfaces of mesosomal tergites I–VI are granular in S. serratus but smooth in S. indus , S. pococki , and S. yaleensis . The dorsosubmedian carinae are granular or costate-granular on metasomal segments I–IV (fig. 76F) in S. serratus , costate on I–IV in S. indus , and costate on I and II or I–III and granular or costate-granular on III and IV or IV, in S. yaleensis . The ventral intercarinal surfaces of metasomal segment IV are smooth in the male and female (fig. 78F) of S. serratus but granular in the male of S. couzijni and S. gravimanus , and the male and female of S. caesar . The dorsolateral carinae of metasomal segment V are distinct and continuous (fig. 77F) in S. serratus but obsolete and discontinuous to absent in S. indus and S. pococki . The dorsal intercarinal surface of metasomal segment V is granular in the male (fig. 76F) but smooth in the female of S. serratus , smooth in the male of S. indus and S. pococki , and granular in the female of S. caesar and S. gravimanus . The telson is dark reddish brown, paler than metasomal segment V, in S. serratus , but blackish, as dark as segment V, in all other species except S. yaleensis . The telson vesicle is globose in S. serratus but elongate in S. caesar , S. couzijni , S. gravimanus , and S. yaleensis .
DISTRIBUTION: This species is endemic to Sri Lanka and has been recorded in the Southern and Uva provinces (fig. 198, table 1). Its distribution appears to overlap those of G. titanicus , S. gravimanus , and S. yaleensis and to be allopatric with the other two species of Srilankametrus occurring on the island.
ECOLOGY: Srilankametrus serratus inhabits deciduous forest on the coastal plains in the south of the island. The locality records for which data are available range from 40 m to 290 m above sea level ( Kovařík et al., 2016). This species is fossorial and pelophilous, excavating singleoccupant, straight, oblique burrows in hard, clayey soils, in earthen banks ( Kovařík et al., 2016). The buthids Isometrus maculatus and Reddyanus loebli were recorded in sympatry.
CONSERVATION STATUS: Srilankametrus serratus is occasionally available in the exotic pet trade.
REMARKS: Srilankametrus serratus was synonymized with S. indus by Couzijn (1981) but revalidated and redescribed by Tikader and Bastawade (1983). Fet (2000) followed Tikader and Bastawade (1983) in regarding S. serratus as a valid species whereas Kovařík (1998, 2004, 2009) followed Couzijn (1981) in treating it as a synonym of S. indus . Recently, Kovařík et al. (2016: 110, 111) redescribed S. serratus based on new material from southern Sri Lanka and confirmed its validity as follows:
Pocock (1900: 86) differentiated H. serratus and H. indus as follows: 1) dorsal and dorsolateral carinae of metasomal segments granulated in H. serratus ... smooth in H. indus .... We found other minor differences: 2) sexual dimorphism in proportions of pedipalps not noticeable in H. indus , male with slightly narrower chela than female in H. serratus (chela length to width ratio 1.79–1.94 in males, 1.69 in the females); 3) carapace smooth and glossy, only occasionally with granules at margins in H. indus ; carapace smooth and glossy medially, always with more granules at margins in H. serratus ... 4) spination formula of tarsomeres II of legs: 3/4: 2–4/3–4: 4/4–5: 4/ 5 in H. indus ; 3–4/4–6: 4/4–5: 4/5–6: 4/ 5–7 in H. serratus .
Based on the present investigation, the species redescribed as S. serratus by Kovařík et al. (2016) is more closely related to S. gravimanus than to S. indus (fig. 10) but can be separated from both species by means of characters recounted in the key and diagnosis. These characters include Pocock’s (1900a) original diagnostic difference in the macrosculpture of the metasomal carinae, as well as the difference in the distribution of carapace surface granulation, i.e., character 3, noted by Kovařík et al. (2016). The sexually dimorphic difference in proportions of the pedipalp chela of the male, i.e., character 2 of Kovařík et al. (2016), supports the closer relationship with S. gravimanus . On the other hand, the counts of telotarsal spiniform macrosetae (character 4) cannot be used to separate these species; the ranges given by Kovařík et al. (2016) for S. indus and S. serratus , as well as S. gravimanus (4/4–5, 4/5, 4/5–6, 4/5–6) overlap completely.
The adult male holotype of S. serratus , redescribed and illustrated by Tikader and Bastawade (1983) and again by Kovařík (2004, 2009), differs from the specimens from southern Sri Lanka, described by Kovařík et al. (2016), as well as material studied for the present investigation, in some respects. Firstly, the carapace shape of the holotype male of S. serratus more closely resembles S. indus , being broader and more markedly dorsoventrally compressed, than other S. serratus material from southern Sri Lanka examined in the present study, and cited by Kovařík et al. (2016), in which the carapace is longer and less dorsoventrally compressed. Secondly, the pedipalp chela manus of the holotype male of S. serratus is not noticeably narrower than the manus of an adult female, a character suggested by Kovařík et al. (2016); on the contrary, it is very broad, more closely resembling the adult male of S. indus . The spiniform macrosetal counts on the telotarsi of legs I–IV of the holotype, incorrectly cited as 5/6, 5/5–6, 5/6, 5/6 by Tikader and Bastawade (1983: 558), but verified as 4/5, 4/5, 4/5, 3–4/ 5 in the present study, fall within the range of the counts of S. serratus material from southern Sri Lanka cited by Kovařík et al. (2016), and four specimens from southern Sri Lanka in the FMNH examined during the present study (3 ♂: 4/4, 4/5, 4/5, 4/5; 4/5, 4/5, 4/5, 4/5; 3/4, 4/4, 4/5, 4/5; 1 subad. ♀: 3/4, 3–4/4, 4/5, 3–4/5) as well as within the range of counts for S. gravimanus and S. indus noted above. Therefore, although the species in southern Sri Lanka is clearly distinct from S. gravimanus and S. indus , it is unclear whether this species is, indeed, conspecific with the holotype of S. serratus , the precise type locality of which is unknown.
MATERIAL EXAMINED: SRI LANKA: Ceylon, E.C. Fernando, 1 ♂, 1 subad. ♀, 1 juv. ♂ (FMNH 086 264), 21.iv.1969, E.C. Fernando, 3 ♂ (FMNH 086 916), 1899, Dr. H. Driesch, 1 ♂, 1 juv. ♂, 1 juv. ♀ (ZMH), x.1912, 1 ♀, 1 juv. ♂ (ZMH ex Mus. Calcutta). Southern Prov.: Galle Distr.: between Bentota and Haburugala, 06°24′N 80°01′E, x.2016, A. Ullrich, 1 juv. ♀ (AMCC [LP 14058]). Uva Prov.: Moneragala Distr.: Warahana [06°27′N 81°27′E], Uva, Ceylon, xi.1962, E.C. Fernando, 1 subad. ♂, 1 subad. ♀ (FMNH 086 251).
Couzijn, H. W. C. 1981. Revision of the genus Heterometrus. Zoologische Verhandelingen 184: 1 - 196.
Fet, V. 2000. Family Scorpionidae Latreille, 1802. In V. Fet, W. D. Sissom, G. Lowe, and M. E. Braunwalder, Catalog of the scorpions of the World (1758 - 1998): 427 - 486. New York: New York Entomological Society.
Kovarik, F. 1998. Stiri [Scorpions]. Jihlava, Czech Republic: Madagaskar, 176 pp. [in Czech]
Kovarik, F. 2004. A review of the genus Heterometrus Ehrenberg, 1828, with descriptions of seven new species (Scorpiones, Scorpionidae). Euscorpius 15: 1 - 60.
Kovarik, F. 2009. Illustrated catalog of scorpions. Part 1. Introductory remarks; keys to families and genera; subfamily Scorpioninae with keys to Heterometrus and Pandinus species. Prague: Clarion Production. 170 pp.
Kraepelin, K. 1899. Scorpiones und Pedipalpi. In F. Dahl (editor), Das Tierreich, 8 (Arachnoidea): 1 - 265. Herausgegeben von der Deutschen Zoologischen Gesellschaft. Berlin: R. Friedlander und Sohn Verlag.
Pliskova, J., F. Kovarik, O. Kosulic, and F. St'ahlavsky. 2016. Description of a new species of Heterometrus Ehrenberg, 1828 (Scorpiones: Scorpionidae) from Thailand with remarks about the utilization of cytogenetic data in taxonomy of the genus. Annales Zoologici (Warszawa) 66: 467 - 476.
Pocock, R. I. 1894 a. A small contribution to our knowledge of the scorpions of India. Annals and Magazine of Natural History (Ser. 6) 13: 72 - 84.
Pocock, R. I. 1900 a. Arachnida. In W. T. Blandford (editor), The fauna of British India, including Ceylon and Burma. London: Taylor and Francis, 279 pp.
Roewer, C. F. 1943. Uber eine neuerworbene Sammlung von Skorpionen des Natur-Museums Senckenberg. Senckenbergiana 26: 205 - 244.
Takashima, H. 1945. [Scorpions of eastern Asia.] Acta Arachnologica 9: 68 - 106. [in Japanese]
Tikader, B. K., and D. B. Bastawade. 1983. Fauna of India, vol. 3. Scorpions (Scorpionida: Arachnida). Zoological Survey of India. Pune: Sangam Press. 671 pp.
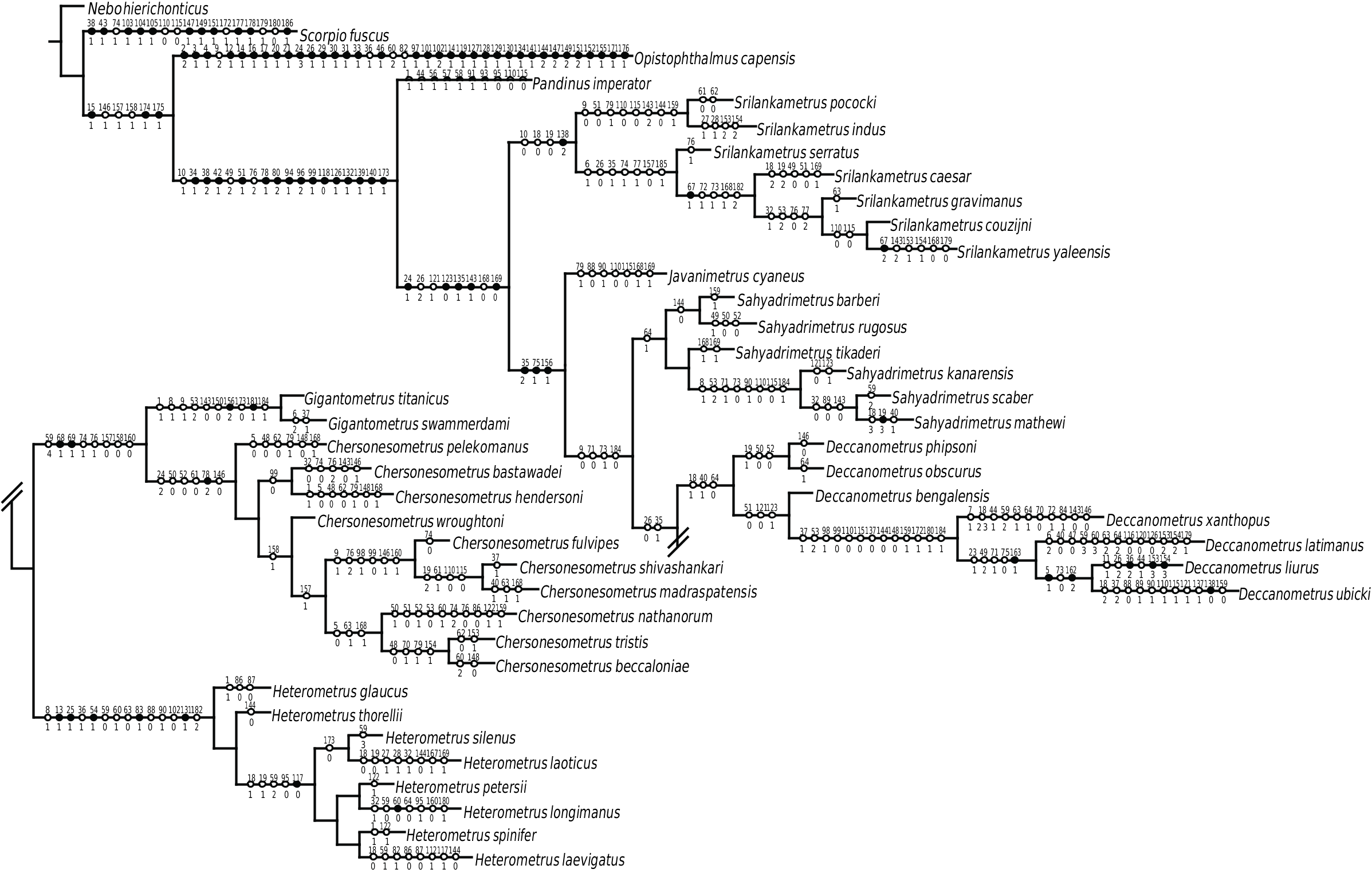
FIGURE 10. Phylogeny of Asian forest scorpions (Scorpionidae: Heterometrinae Simon, 1879, stat. nov.), obtained by simultaneous phylogenetic analysis of 186 morphological characters and 4188 aligned DNA nucleotides from three mitochondrial and two nuclear gene loci (Loria and Prendini, in press). Maximum likelihood tree with unambiguous morphological synapomorphies optimized. Black circles indicate uniquely derived apomorphic states, white circles parallel derivations of apomorphic states. Numbers above circles indicate characters, numbers below indicate states (appendix 1).
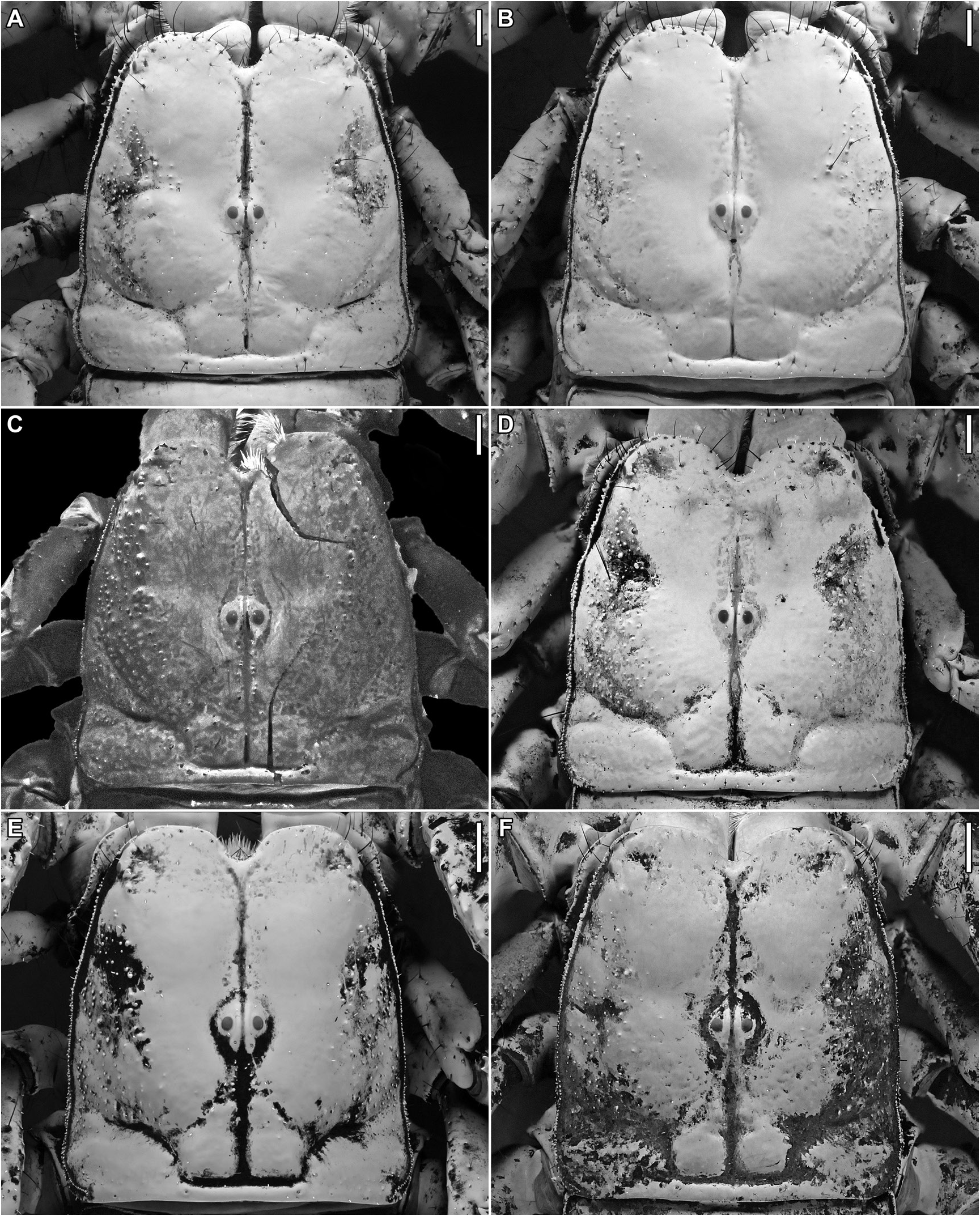
FIGURE 28. Srilankametrus Couzijn, 1981, stat. nov. et stat. rev., carapace, dorsal aspect. A, B. Srilankametrus indus (DeGeer, 1778), comb. nov., A. ♂, B. ♀ (AMNH), Kandy, Sri Lanka. C, E, F. Srilankametrus serratus (Pocock, 1900), comb. nov., C. holotype ♂ (BMNH 1888.55), E. ♂ (FMNH 086 916), F. ♀ (ZMH), Ceylon [Sri Lanka]. D. Srilankametrus pococki, sp. nov., holotype ♀ (AMNH), Kodigala, Sri Lanka. Scale bars = 2 mm.
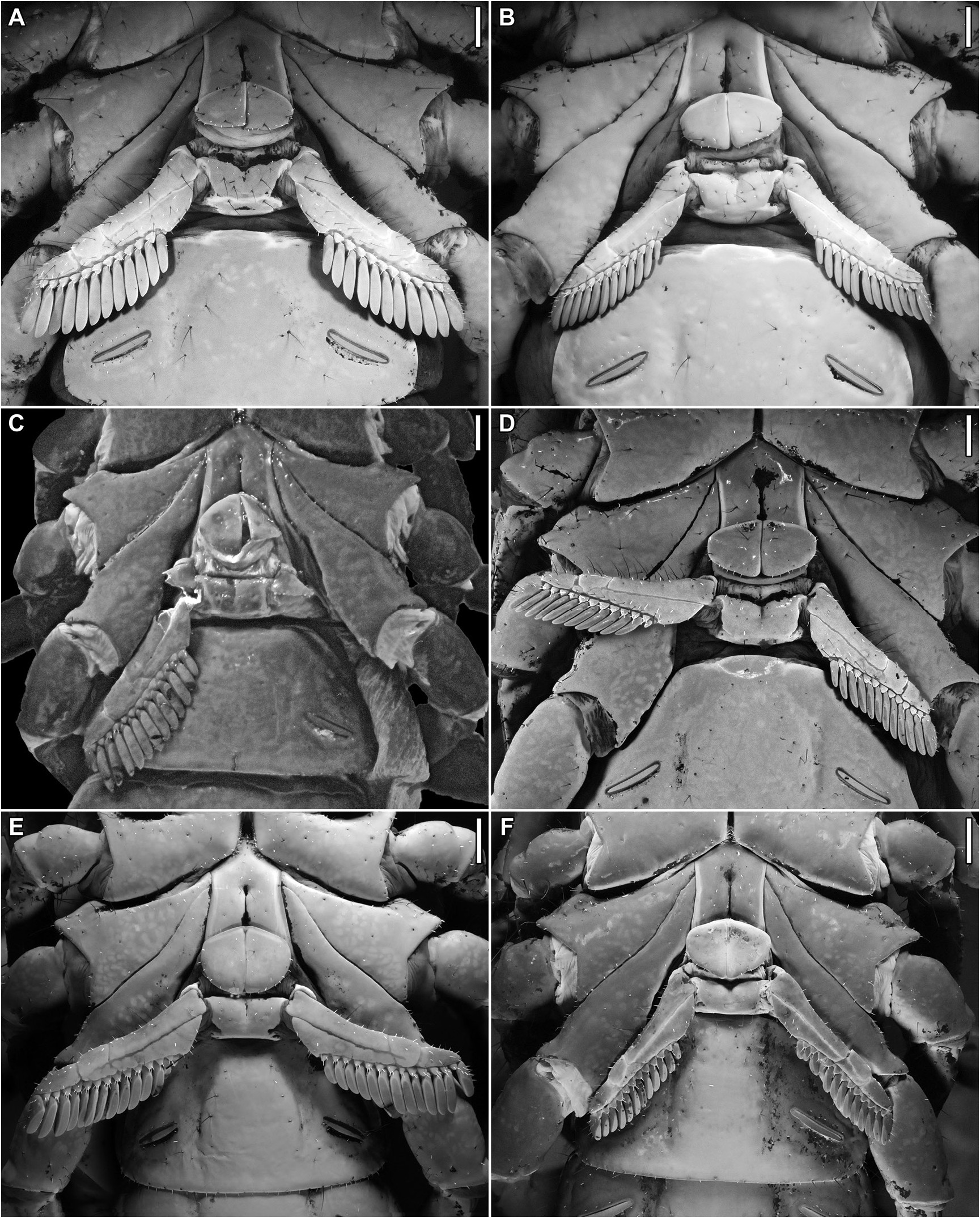
FIGURE 42. Srilankametrus Couzijn, 1981, stat. nov. et stat. rev., sternum and pectines, ventral aspect. A, B. Srilankametrus indus (DeGeer, 1778), comb. nov., A. ♂, B. ♀ (AMNH), Kandy, Sri Lanka. C, E, F. Srilankametrus serratus (Pocock, 1900), comb. nov., C. holotype ♂ (BMNH 1888.55), E. ♂ (FMNH 086 916), F. ♀ (ZMH), Ceylon [Sri Lanka]. D. Srilankametrus pococki, sp. nov., holotype ♀ (AMNH), Kodigala, Sri Lanka. Scale bars = 2 mm.

FIGURE 57. Srilankametrus Couzijn, 1981, stat. nov. et stat. rev., legs I–IV, tibiae, basitarsi and telotarsi, ventral aspect. A–D. Srilankametrus indus (DeGeer, 1778), comb. nov., ♂ (AMNH), Kandy, Sri Lanka. E–H. Srilankametrus pococki, sp. nov., holotype ♀ (AMNH), Kodigala, Sri Lanka. I–L. Srilankametrus serratus (Pocock, 1900), comb. nov., ♂ (ZMH), Ceylon [Sri Lanka]. Scale bars = 2 mm.

FIGURE 76. Srilankametrus Couzijn, 1978, stat. nov. et stat. rev., metasoma and telson, dorsal aspect. A. Srilankametrus caesar (C.L. Koch, 1841), comb. nov. et stat. rev., ♂ (MNHN RS 0079). B. Srilankametrus couzijni, sp. nov., holotype ♂ (BMNH 1896.7.30.121). C. Srilankametrus gravimanus (Pocock, 1894), comb. nov., ♂ (AMNH), Kokmotte, Sri Lanka. D. Srilankametrus indus (DeGeer, 1778), comb. nov., ♂ (AMNH), Kandy, Sri Lanka. E. Srilankametrus pococki, sp. nov., holotype ♀ (AMNH), Kodigala, Sri Lanka. F. Srilankametrus serratus (Pocock, 1900), comb. nov., ♂ (ZMH), Ceylon [Sri Lanka]. Scale bars = 5 mm

FIGURE 77. Srilankametrus Couzijn, 1978, stat. nov. et stat. rev., metasoma and telson, lateral aspect. A. Srilankametrus caesar (C.L. Koch, 1841), comb. nov. et stat. rev., ♂ (MNHN RS 0079). B. Srilankametrus couzijni, sp. nov., holotype ♂ (BMNH 1896.7.30.121). C. Srilankametrus gravimanus (Pocock, 1894), comb. nov., ♂ (AMNH), Kokmotte, Sri Lanka. D. Srilankametrus indus (DeGeer, 1778), comb. nov., ♂ (AMNH), Kandy, Sri Lanka. E. Srilankametrus pococki, sp. nov., holotype ♀ (AMNH), Kodigala, Sri Lanka. F. Srilankametrus serratus (Pocock, 1900), comb. nov., ♂ (ZMH), Ceylon [Sri Lanka]. Scale bars = 5 mm.

FIGURE 78. Srilankametrus Couzijn, 1978, stat. nov. et stat. rev., metasoma and telson, ventral aspect. A. Srilankametrus caesar (C.L. Koch, 1841), comb. nov. et stat. rev., ♂ (MNHN RS 0079). B. Srilankametrus couzijni, sp. nov., holotype ♂ (BMNH 1896.7.30.121). C. Srilankametrus gravimanus (Pocock, 1894), comb. nov., ♂ (AMNH), Kokmotte, Sri Lanka. D. Srilankametrus indus (DeGeer, 1778), comb. nov., ♂ (AMNH), Kandy, Sri Lanka. E. Srilankametrus pococki, sp. nov., holotype ♀ (AMNH), Kodigala, Sri Lanka. F. Srilankametrus serratus (Pocock, 1900), comb. nov., ♂ (ZMH), Ceylon [Sri Lanka]. Scale bars = 5 mm.
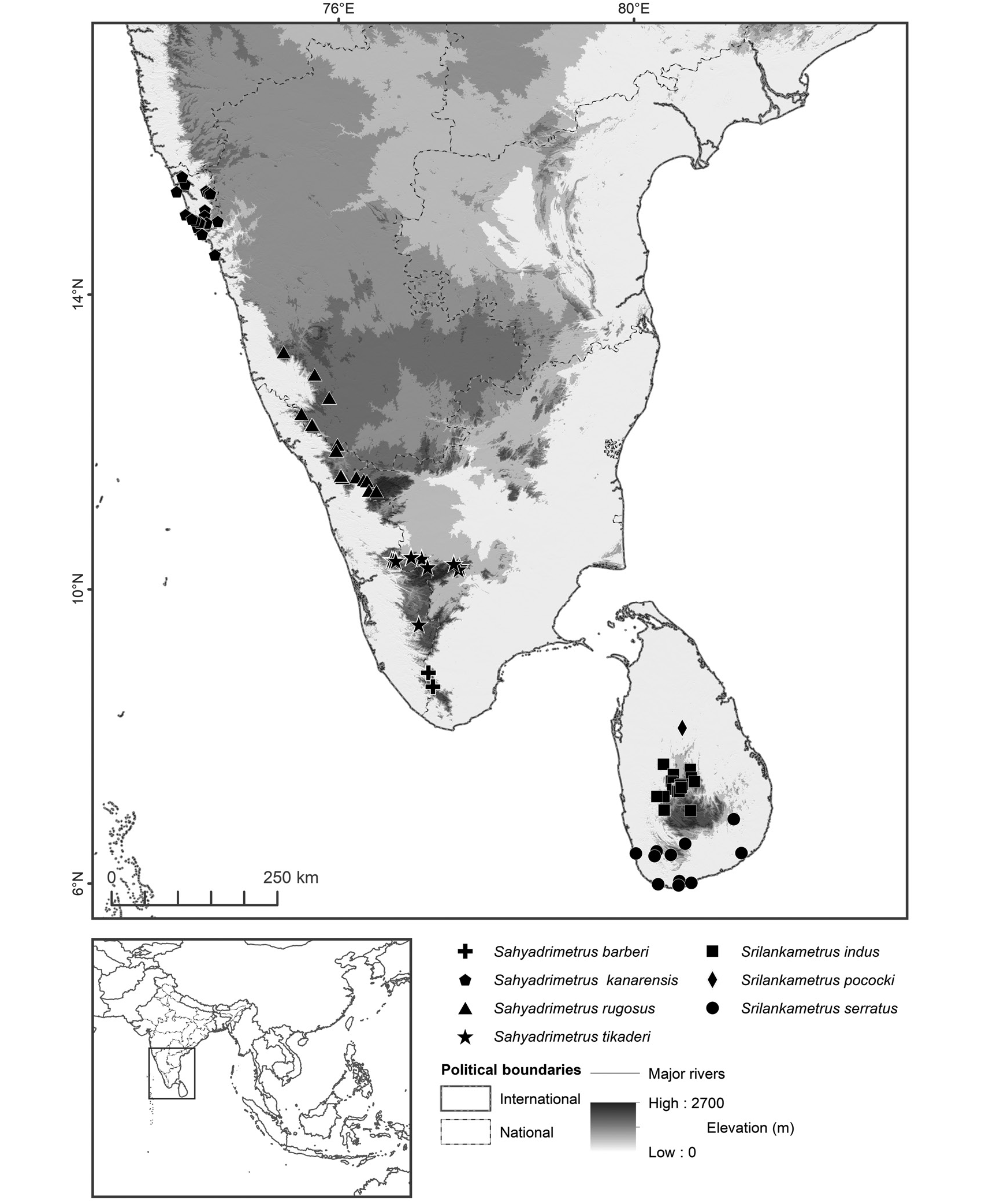
FIGURE 198. Map of South India and Sri Lanka, plotting known locality records of four species of Sahyadrimetrus, gen. nov., and three species of Srilankametrus Couzijn, 1981, stat. nov. et stat. rev., based on material examined, and verified records from virtual museums and the literature: Sahyadrimetrus barberi (Pocock, 1900), comb. nov.; Sahyadrimetrus kanarensis (Pocock, 1900), comb. nov.; Sahyadrimetrus rugosus (Couzijn, 1981), comb. et stat. nov.; Sahyadrimetrus tikaderi, gen. et sp. nov.; Srilankametrus indus (DeGeer, 1778), comb. nov.; Srilankametrus pococki, sp. nov.; Srilankametrus serratus (Pocock, 1900), comb. nov.

FIGURE 244. Srilankametrus serratus (Pocock, 1900), comb. nov., habitus: A, C. dorsal and B, D. ventral aspects. A, B. ♂ (ZMH), C, D. ♀ (ZMH), Ceylon [Sri Lanka]. Scale bars = 10 mm.
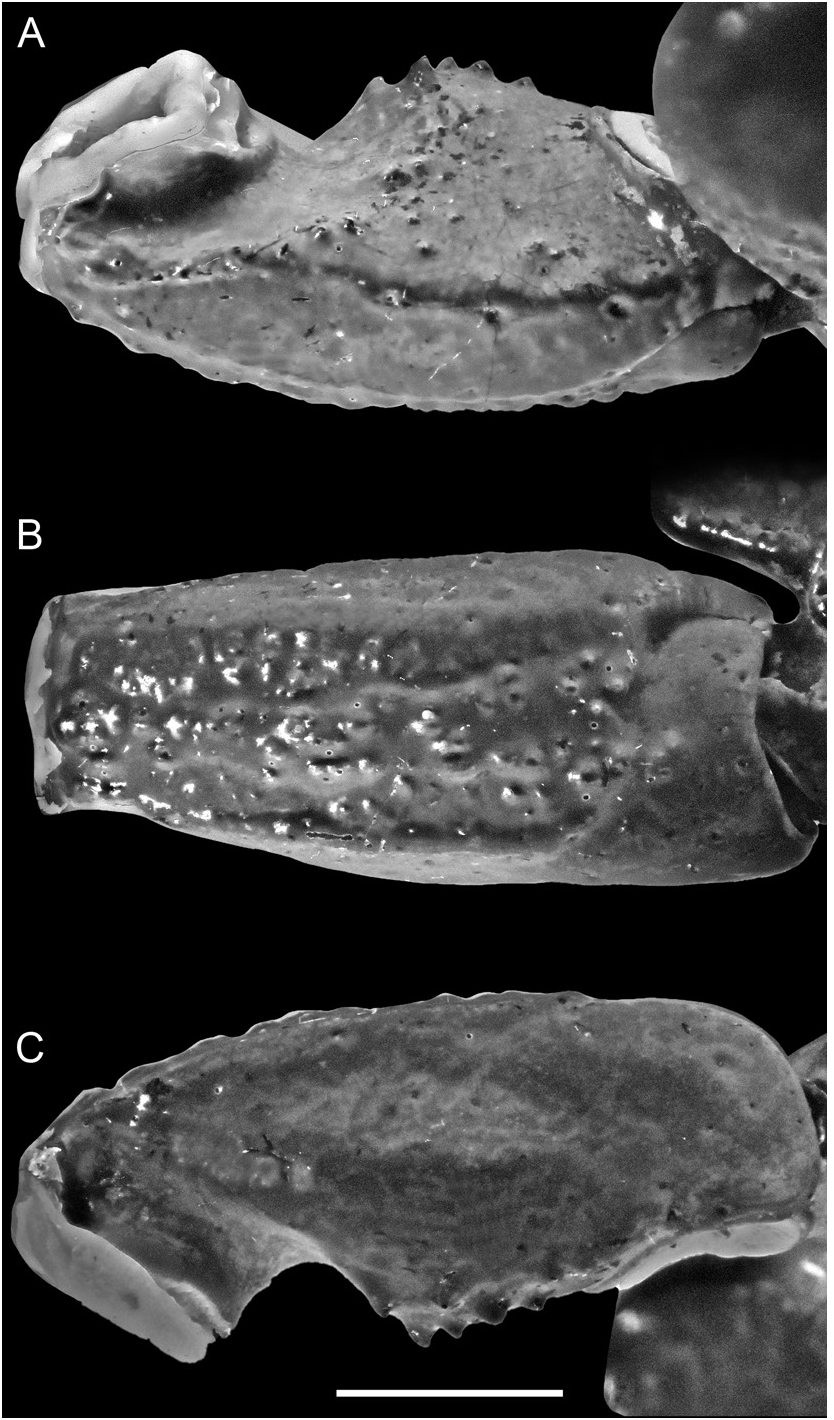
FIGURE 247. Srilankametrus serratus (Pocock, 1900), comb. nov., holotype ♂ (BMNH 1888.55), pedipalp patella: A. dorsal, B. retrolateral, and C. ventral aspects. Scale bar = 5 mm.

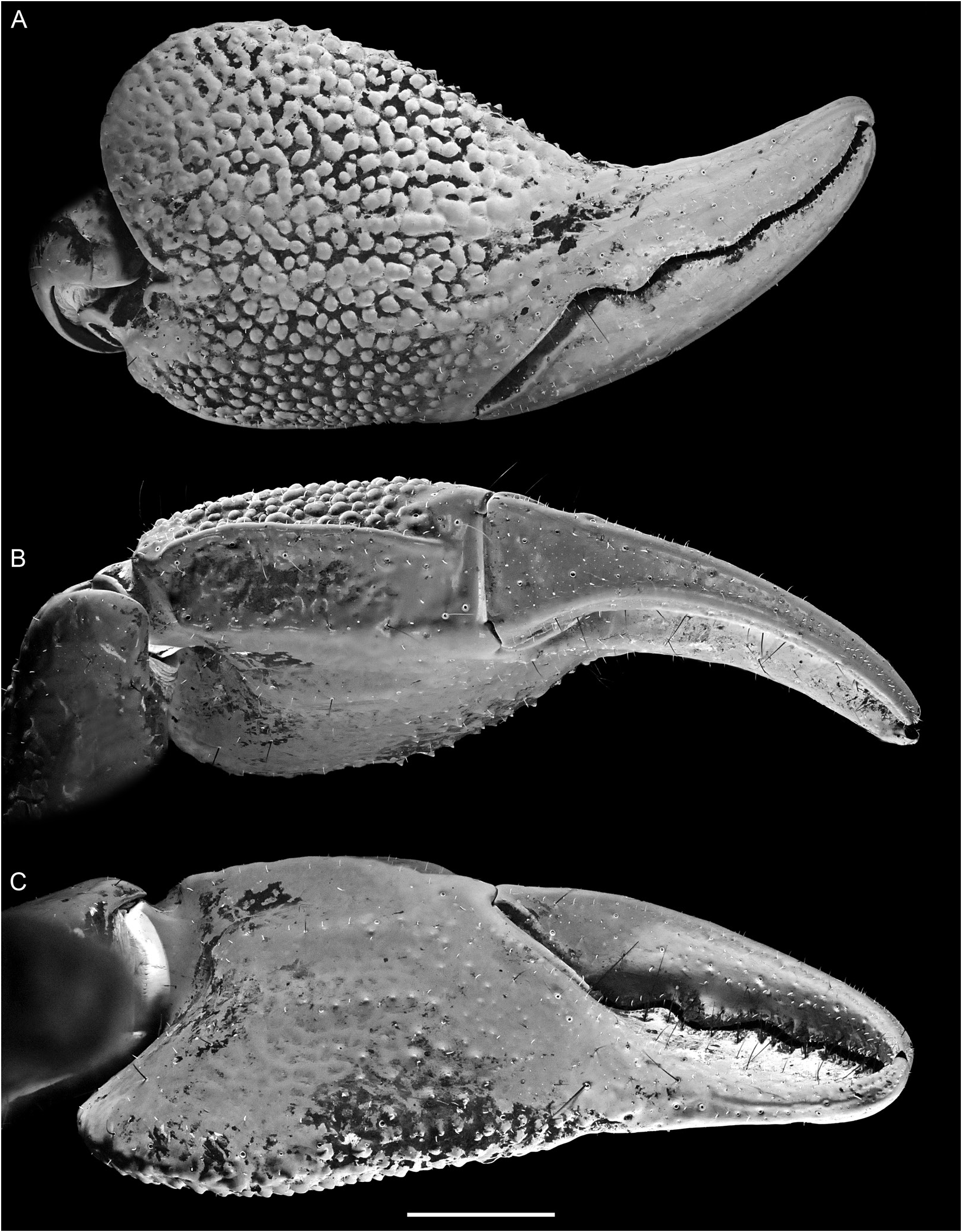
FIGURE 248. Srilankametrus serratus (Pocock, 1900), comb. nov., holotype ♂ (BMNH 1888.55), pedipalp chela: A. retrodorsal, B. ventral, and C. prolateral aspects. Scale bar = 5 mm.

FIGURE 249. Srilankametrus serratus (Pocock, 1900), comb. nov., ♂ (FMNH 086 916), pedipalp patella: A. dorsal, B. retrolateral, and C. ventral aspects. Scale bar = 5 mm.

FIGURE 250. Srilankametrus serratus (Pocock, 1900), comb. nov., ♂ (FMNH 086 916), pedipalp chela: A. retrodorsal, B. ventral, and C. prolateral aspects. Scale bar = 5 mm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Srilankametrus serratus (Pocock, 1900)
| Prendini, Lorenzo & Loria, Stephanie F. 2020 |
Heterometrus (Srilankametrus) serratus:
| Fet, V. 2000: 448 |
| Tikader, B. K. & D. B. Bastawade 1983: 545 |
Heterometrus (Srilankametrus) indus: Couzijn, 1981: 82
| Fet, V. 2000: 446 |
| Kovarik, F. 1998: 137 |
| Couzijn, H. W. C. 1981: 82 |
Heterometrus (Srilankametrus) indus indus: Couzijn, 1981: 39
| Fet, V. 2000: 447 |
| Couzijn, H. W. C. 1981: 39 |
Heterometrus serratus:
| Takashima, H. 1945: 94 |
Palamnaeus caesar: Pocock, 1900a: 86
| Pocock, R. I. 1900: 86 |
Palamnaeus serratus
| Kovarik, F. 2004: 17 |
| Pocock, R. I. 1900: 86 |
Heterometrus caesar: Kraepelin, 1899: 109
| Takashima, H. 1945: 92 |
| Roewer, C. F. 1943: 229 |
| Kraepelin, K. 1899: 109 |
Scorpio caesar:
| Pocock, R. I. 1894: 73 |