
Guimaraesiella ( Cicchinella ) corrugata, Gustafsson & Tian & Ren & Liu & Yu & Zou, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.5060.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:A9780904-16FA-4006-A8DE-E301D40294F0 |
|
DOI |
https://doi.org/10.5281/zenodo.5636400 |
|
persistent identifier |
https://treatment.plazi.org/id/617087CC-6B6D-E25B-C0BE-FCD8E1E5FF4A |
|
treatment provided by |
Plazi |
|
scientific name |
Guimaraesiella ( Cicchinella ) corrugata |
| status |
sp. nov. |
Guimaraesiella ( Cicchinella) corrugata new species
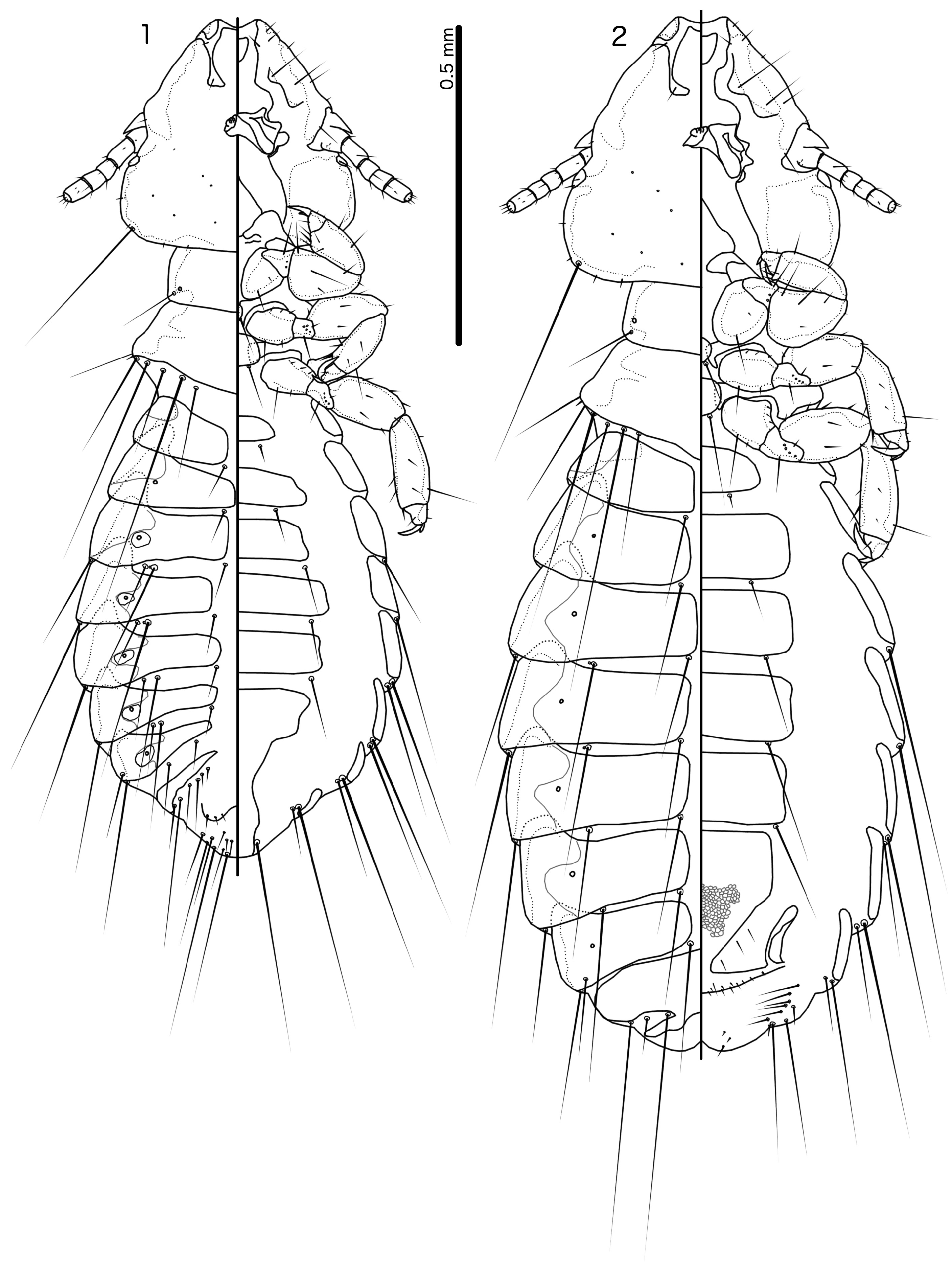
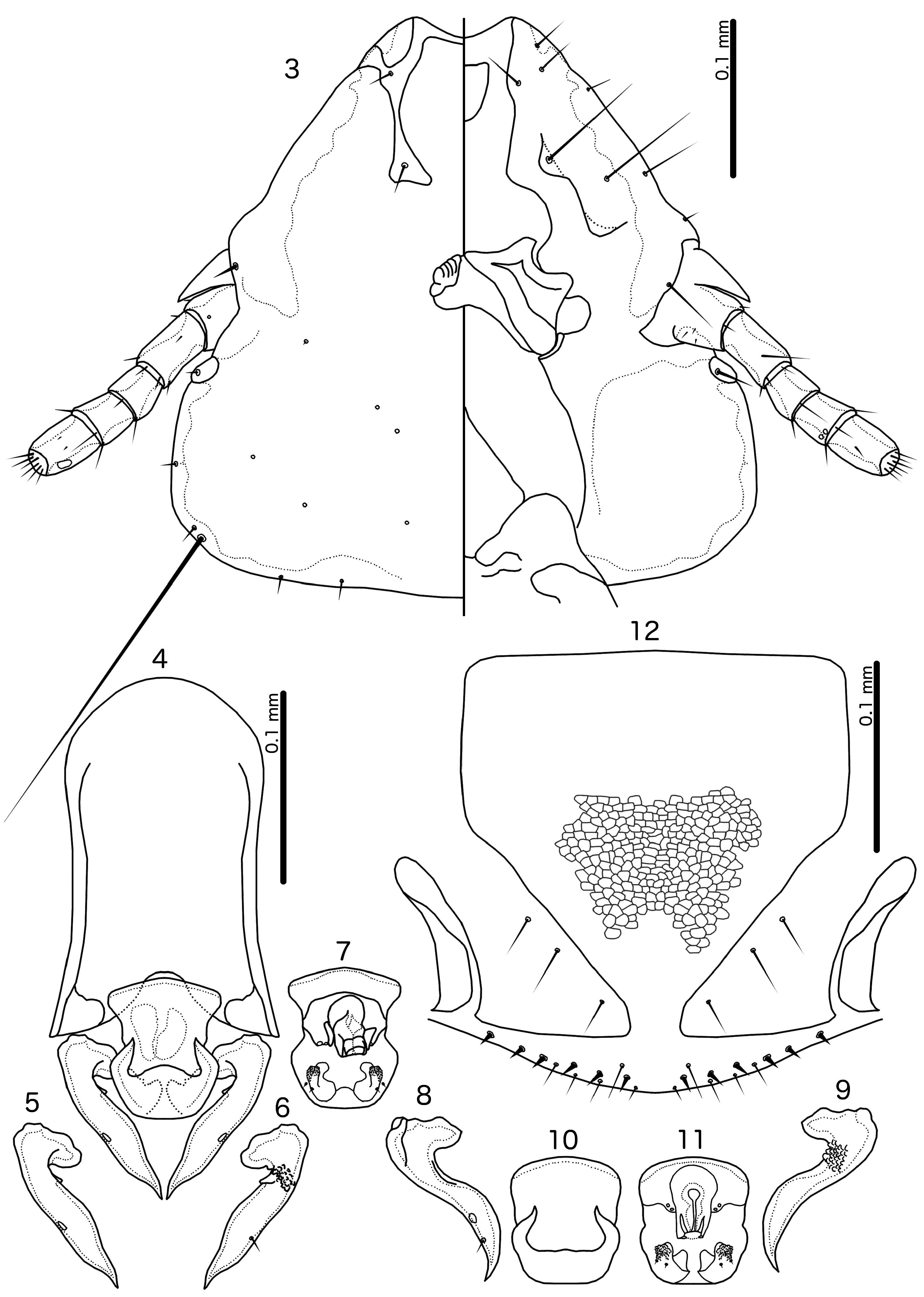
( Figs 1–12 View FIGURES 1–2 View FIGURES 3–12 )
Type host. Alcippe hueti hueti David, 1874 – Huet’s fulvetta ( Leiothrichidae ) [see Remarks].
Type locality. Dinghushan , Dinghu District, Zhaoqing, Guangdong, China .
gnosis. In the key to species in Gustafsson et al. (2019a), Guimaraesiella ( Cicchinella) corrugata keys out to Guimaraesiella ( Cicchinella) mcgrewi Gustafsson et al., 2019a based on the extent of the dorsal preantennal suture, the structure of the male genitalia, and the fusion of female tergopleurites IX+X and XI. However, these two species can be separated by the following characters: lateral margins of preantennal head more convex in G. ( C.) corrugata ( Fig. 3 View FIGURES 3–12 ) than in G. ( C.) mcgrewi ; parameral heads with single-pointed median ends of the median extensions in G. ( C.) corrugata ( Figs 5, 8 View FIGURES 3–12 ), but with double-pointed median ends in G. ( C.) mcgrewi ; gonopore small with triangular lateral extensions in G. ( C.) mcgrewi , but large and complicated, without triangular extensions but with large, rough areas bearing what appears to be 1–2 sensilla on each side in G. ( C.) corrugata ( Figs 7, 11 View FIGURES 3–12 ); rugose nodi submarginal in G. ( C.) corrugata ( Figs 7, 11 View FIGURES 3–12 ), but marginal in G. ( C.) mcgrewi ; patterns of thickenings of distal mesosome in G. ( C.) corrugata ( Figs 4, 7, 10, 11 View FIGURES 3–12 ) different from those of G. ( C.) mcgrewi ; male tergopleurite VIII with two tps on each side in G. ( C.) mcgrewi , but with 1 tps on each side in G. ( C.) corrugata ( Fig. 1 View FIGURES 1–2 ); female subgenital plate with extensive reticulation in G. ( C.) corrugata ( Fig. 12 View FIGURES 3–12 ), but without reticulation in G. ( C.) mcgrewi .
Description. Both sexes. Head sub-triangular, frons concave ( Fig. 3 View FIGURES 3–12 ). Marginal carina interrupted laterally and medianly; width of carina irregular, widest near preantennal nodi. Dorsal preantennal suture reaches lateral margins of head and ads, but does not cut off dorsal anterior plate medianly. Head chaetotaxy as in Fig, 3; pos situated ventrally in male, more dorsally in female. Preantennal nodi large, bulging, somewhat pointed medianly, but shape variable among specimens. Antennae not sexually dimorphic or with scape only slightly stouter in male. Preocular nodi much larger than postocular nodi. Temporal marginal carina narrow, with undulating median margin. Gular plate pointed anteriorly. Thoracic and abdominal segments as in Figs 1–2 View FIGURES 1–2 . Measurements as in Table 1 View TABLE 1 .
Male. Thoracic and abdominal chaetotaxy as in Fig. 1 View FIGURES 1–2 . One tps on each side of tergopleurite VII. The genitalia differ between two populations: specimens from Dinghushan (including holotype) as in Figs 4–7 View FIGURES 3–12 , and those from Dadongshan with parameres as in Figs 8–9 View FIGURES 3–12 and mesosome as in Figs 10–11 View FIGURES 3–12 . Mesosome longer than wide, with rounded proximal end protruding laterally (Dinghushan specimens, Fig. 7 View FIGURES 3–12 ); lateral margins deeply concave at about mid-section in specimens from Dinghushan ( Fig. 7 View FIGURES 3–12 ), but almost straight in specimens from Dadongshan ( Figs 10–11 View FIGURES 3–12 ); distal margin of mesosome flat or slightly convex, with wide dorsal thickening, curved at lateral ends ( Figs 4, 10 View FIGURES 3–12 ). Mesosome with a ventral, distal pair of submarginal elongated nodi associated with prominent rugose areas on antero-lateral ends on both sides, and 2 pmes microsetae on each side of elongated nodi ( Figs 7, 11 View FIGURES 3–12 ). Gonopore large, tube-like, arising in anterior end of mesosome; 1–2 ames sensilla on more or less protruding areas on each side of gonopore. Parameres with pointed heads ( Figs 5–6, 8–9 View FIGURES 3–12 ), ventrally with densely corrugated area on the distal base of head. Parameral blades bulky, curved in specimens from Dadongshan ( Figs 8–9 View FIGURES 3–12 ) but more straight in specimens from Dinghushan ( Figs 5–6 View FIGURES 3–12 ); pst1–2 as in Figs 5–6 and 8 View FIGURES 3–12 ; pst1 positioned ventrally in specimens from Dinghushan, but dorsally in specimens from Dadongshan. However, in both cases pst2 seta is situated near the lateral margin, and may in fact be marginal in unmounted specimens.
Female. Thoracic and abdominal chaetotaxy as in Fig. 2 View FIGURES 1–2 . Subgenital plate rectangular anteriorly, with distal part gently narrowing to slender connection with complete cross-piece ( Fig. 12 View FIGURES 3–12 ). Irregular reticulation present in distal half of subgenital plate, less prominent in more anterior parts as in Fig. 12 View FIGURES 3–12 . Vulval margin with 4 short, slender vms and 6 short, thorn-like vss on each side; 4 short, slender vos on each side of subgenital plate; distal 1 vos one each side median to or slightly anterior to row of vss.
Etymology. The species epithet derived from corrugatus, Latin for “wrinkled”, referring to the parameral heads.
Type material. Ex Alcippe hueti hueti : Holotype ♂, Dinghushan , Dinghu District, Zhaoqing, Guangdong, China, 18 March 2019, D.R. Gustafsson, L. Lei, C. Adam, G. Chisamera, Bird-ID J4116, Louse-ID GD-PHTH- 00265 ( IZGAS) . Paratypes. 1♂, same data as holotype, Louse-ID GD-PHTH-00266 ( IZGAS) . 2♂, 2♀, same locality and collectors as holotype, 17 Mar. 2019, Bird-ID J4109, Louse-ID GD-PHTH-00267–70 ( IZGAS) .
Non-types: 2♂, 2♀, Dadongshan , Nanling Mountains, Guangdong, China, 11 Mar. 2019, D.R. Gustafsson, L. Lei, C. Adam, G. Chisamera, Bird-ID J4077, Louse-ID GD-PHTH-00271–4 ( IZGAS) .
Remarks. The six males examined differ between the two localities in overall size, particularly in the head, with specimens from Dadongshan being bigger than specimens from Dinghushan. Also, the shape of the male genitalia is different between specimens from the two localities, in particular the mesosome and parameres, which are illustrated separately from each population ( Figs 4–7 and 8–11 View FIGURES 3–12 ). The distal parameres of the male genitalia are partially everted, approximately to the same degree, in the two illustrated specimens; therefore, differences in their morphology are unlikely to be due to artifacts of position. However, the asymmetry of the gonopore in the Dinghushan specimens may be due to slide mounting, and this structure is likely more similar between specimens from the two populations.
It is unclear whether differences in the shape of the mesosome and parameres are taxonomically significant. Since the two collection localities are placed on different mountain ranges, it is possible that the lice on Alcippe hueti hueti differ between ranges, even if the host species and subspecies are the same. More collections in the mountain ranges of South China are needed to establish whether the louse populations from different ranges are effectively isolated from each other, even if the hosts may not be.
Moreover, at the time of collection, we had reason to suspect that some of the birds caught at Dadongshan may have originated from further north, as they were morphologically different (DRG pers. obs.). Species of Alcippe in South China form a complex of closely related and morphologically similar taxa. It is possible that some of the hosts at Dadongshan belonged to Alcippe davidi Styan, 1896 , the species living in central China, north of the Nanling Mountains; however, this could not be established in the field. Since the specimens of Guimaraesiella described here were taken from birds identified as A. hueti hueti in the field at Dinghushan, this host and locality are regarded as type host and type locality of G. ( C.) corrugata . However, if more than one species of Alcippe is regularly present in Dadongshan, at least during the winter, it is conceivable that G. ( C.) corrugata may occur on both host species. More collections from birds on the north side of the Nanling Mountains, including birds positively identified as A. davidi , are needed to confirm this hypothesis.
Notably, intraspecific differences in size appear to be common among lice belonging to the Brueelia -complex. Gustafsson & Bush (2017: 66) noted that, in general, head dimensions were smaller in specimens of Acronirmus gracilis ( Burmeister, 1838) from southern localities than specimens more northern localities, and that lice from migratory swallows collectively contained both small-headed and large-headed lice. Also, Gustafsson & Bush (2017: 172) noted that heads of Traihoriella binhchauensis (Najer & Sychra [in Najer et al.], 2014) showed the same pattern, lice from more southern localities had smaller heads than those from more northern localities. A similar pattern is also seen in a poorly differentiated complex of species in the genus Sturnidoecus Eichler, 1944 ( Gustafsson & Bush 2017: 234). Collectively, these examples suggest that differences in size alone may not be a good indicator of species limits in the Brueelia -complex.
Furthermore, Gustafsson & Bush (2017: 267) noted that Southeast Asian populations of Rostrinirmus ruficeps (Nitzsch [in Giebel], 1866) were morphologically identical to populations from other parts of Eurasia, except for significant differences in the male genitalia. In particular, mesosomes of Southeast Asian lice are asymmetrical, whereas those from other parts of Eurasia are symmetrical. As most species in the Brueelia -complex are known from only one or a few localities, even in cases where more than one host specimen has been examined, the extent of such male genitalic differences occurring among louse populations in different parts of the host’s range is unknown.
| IZGAS |
Georgian Academy of Sciences, Insititute of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Cicchinella |