
Ptychogaster ( Temnoclemmys ) cf. bardenensis Murelaga, Lapparent de Broin, Pereda Suberbiola & Astibia, 1999
|
publication ID |
https://doi.org/10.5252/geodiversitas2021v43a3 |
|
publication LSID |
urn:lsid:zoobank.org:pub:AA28648F-709F-44D6-A3CF-0C1D78B971CF |
|
DOI |
https://doi.org/10.5281/zenodo.4548640 |
|
persistent identifier |
https://treatment.plazi.org/id/6504B17A-FFBD-654B-FCF7-C662F0732A64 |
|
treatment provided by |
Felipe |
|
scientific name |
Ptychogaster ( Temnoclemmys ) cf. bardenensis Murelaga, Lapparent de Broin, Pereda Suberbiola & Astibia, 1999 |
| status |
|
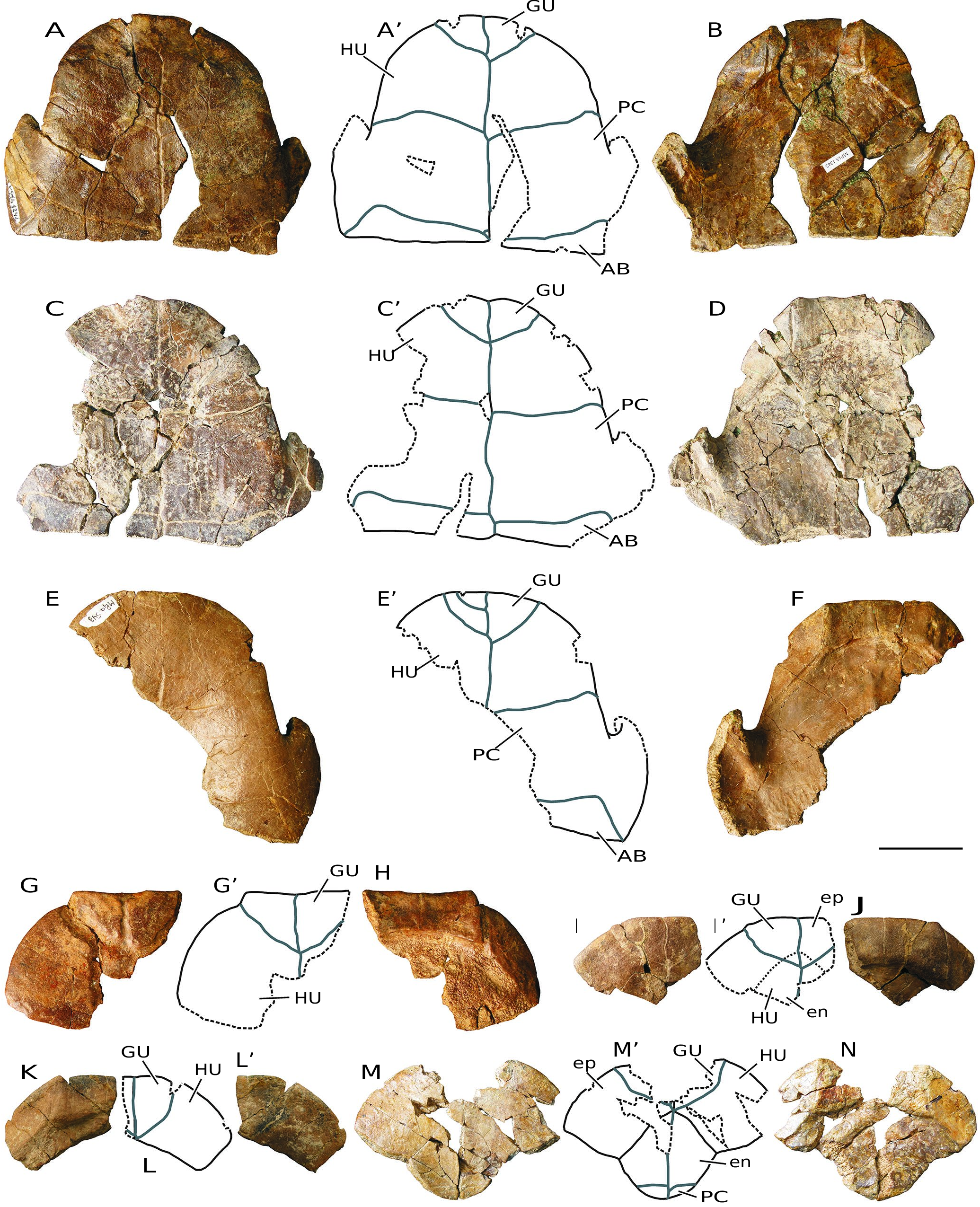
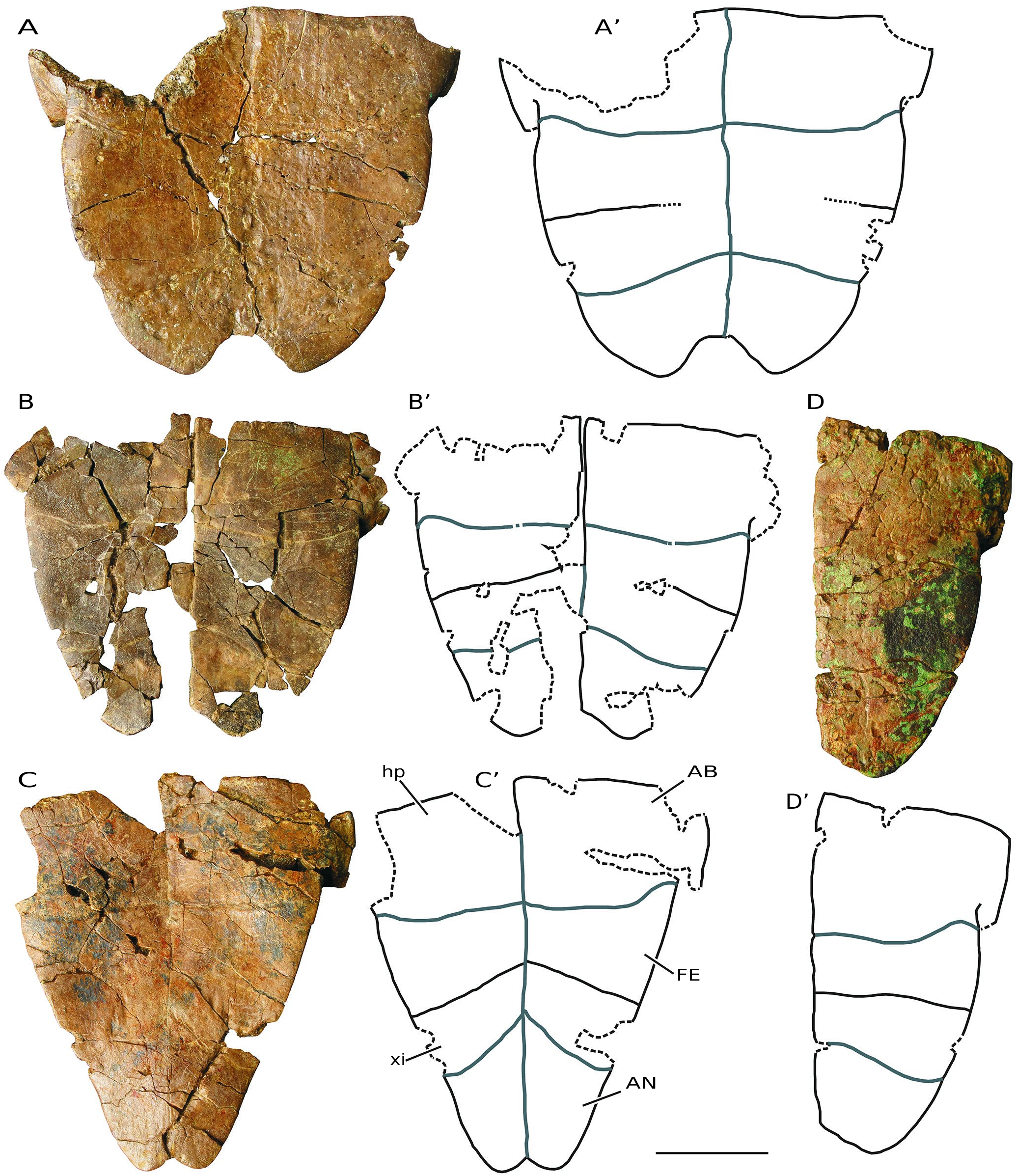
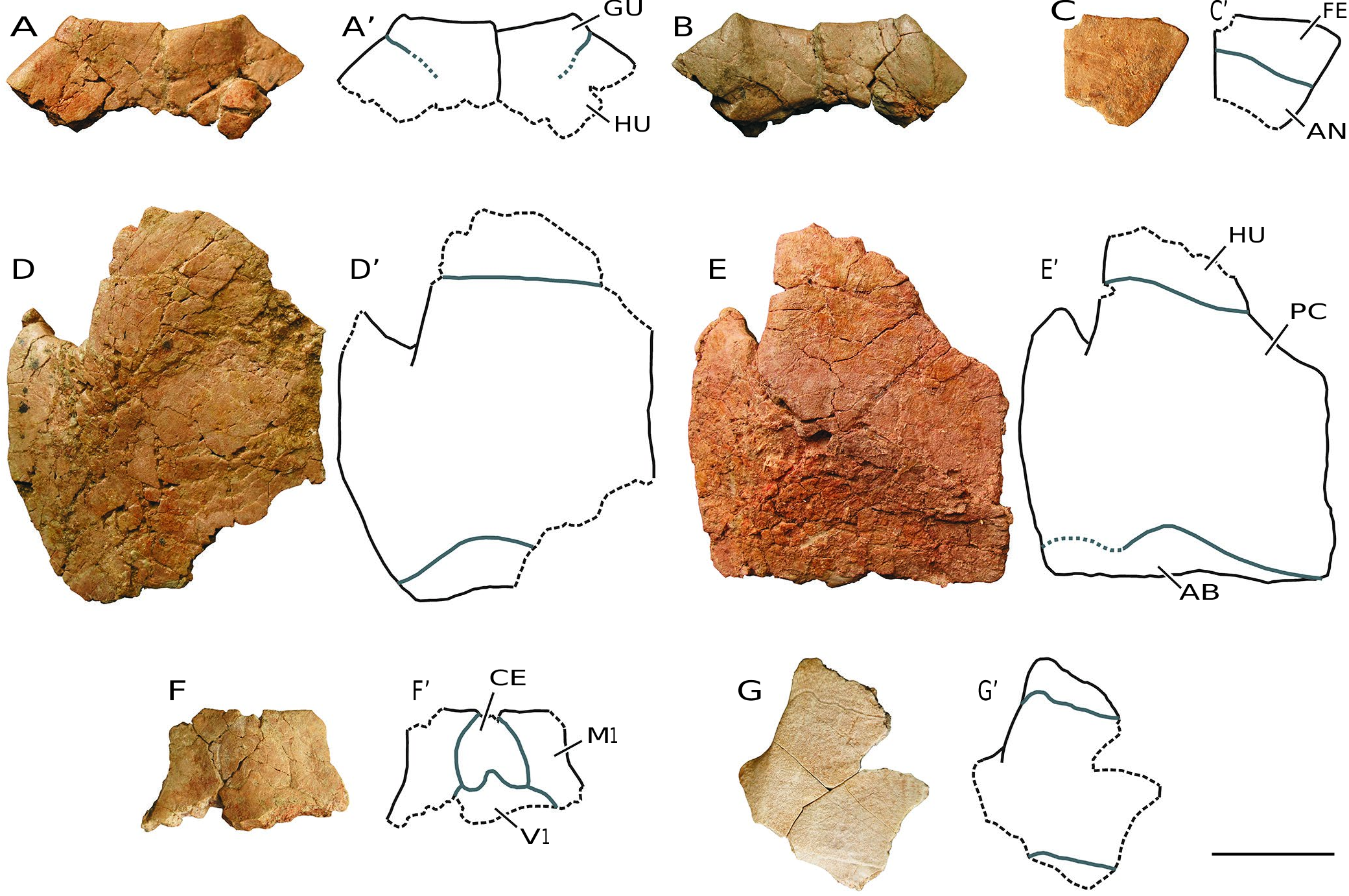
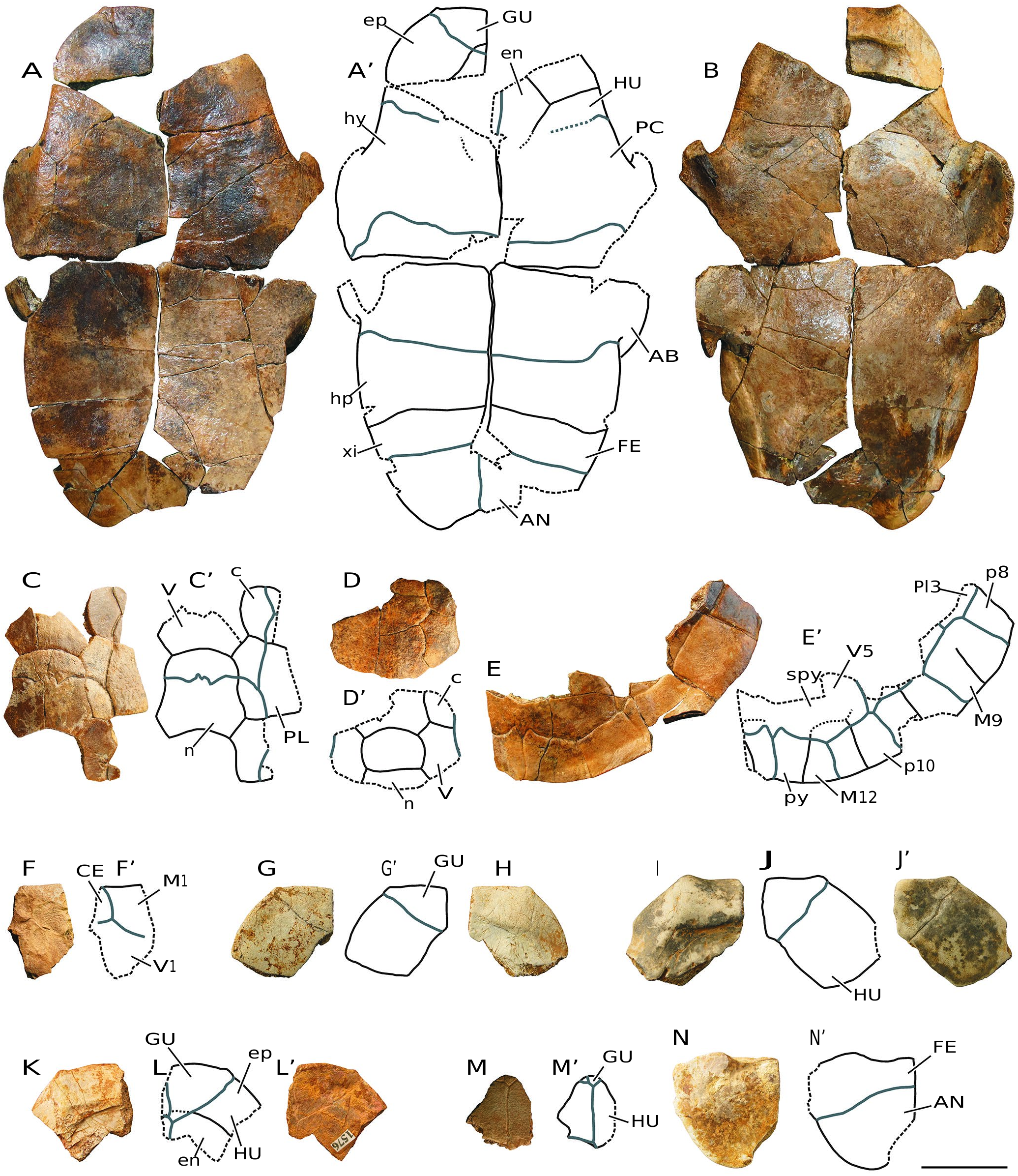
( Figs 3-10 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
MATERIAL EXAMINED. — Numerous disjointed and isolated plates, several partial carapace and plastra, and some relatively complete shells from Monte Pila (e.g., MP 1a-6, MP 1a-14, MP 1a-48, MP 1a- 55, MP 1a-56, MP 1a-81, MP 1a-109, MP 1a-169, MP 1a-180a, MP 1a-228, MP 1a-252, MP 1a-263, MP 1a-277a, MP 1a-281, MP 1a-341, MP 1a-347a, MP 1a-367, MP 1b-369, MP 1b-380b, MP 1a-385, MP 1a-409, MP 1a-449, MP 1a-474, MP 1a-475, MP 1a- 511, MP 1a-528, MP 1a-529b, MP 1a-543, MP 1a-638, MP 1a-643, MP 1a-659, MP 1a-672, MP 1a-679, MP 1a-681, MP 1a-694, MP 1a- 707, MP 1a-757, MP 1a-865, MP 1a-879, MP 1a-911, MP 1a-918, MP 1a-975a, MP 1a-1025, MP 1a-1066, MP 1a-1068b, MP 1a-1070, MP 1a-1145a, MP 1a-1189a, MP 1a-1194, MP 1a-1210a, MP 1a-1221, MP 1a-1227, MP 1a-1242, MP 1a-1305, MP 1a-1333, MP 1a-1342, MP 1a-1345, MP 1a-1350, MP 1a-1364, MP 1a-1394b, MP 1a-1417, MP 1a-1421, MP 1a-1422, MP 1a-1423, MP 1a-1557, MP 1a-1570, MP 1a-1594, MP 1a-1679b, MP 1a-1724, MP 1a-1745, MP 1b-95, MP 1b-169, MP 1b-415, MP 1b-456, MP 1b-467, MP 1b-481, MP 1b-556, MP 1b-585, MP 1b-1597a, MP 1b-sup1-a, MP 2-93, MP 2-175, MP 2-306, MP 2-307, MP 2-308, MP 3-1; Figs 3-9 View FIG View FIG View FIG View FIG View FIG View FIG View FIG ) and Fuenmayor (e.g., FM1-1, FM1-2, FM1-3, FM1-4, FM2-1, MCNA 16032, MCNA 16031; Fig. 10 View FIG ).
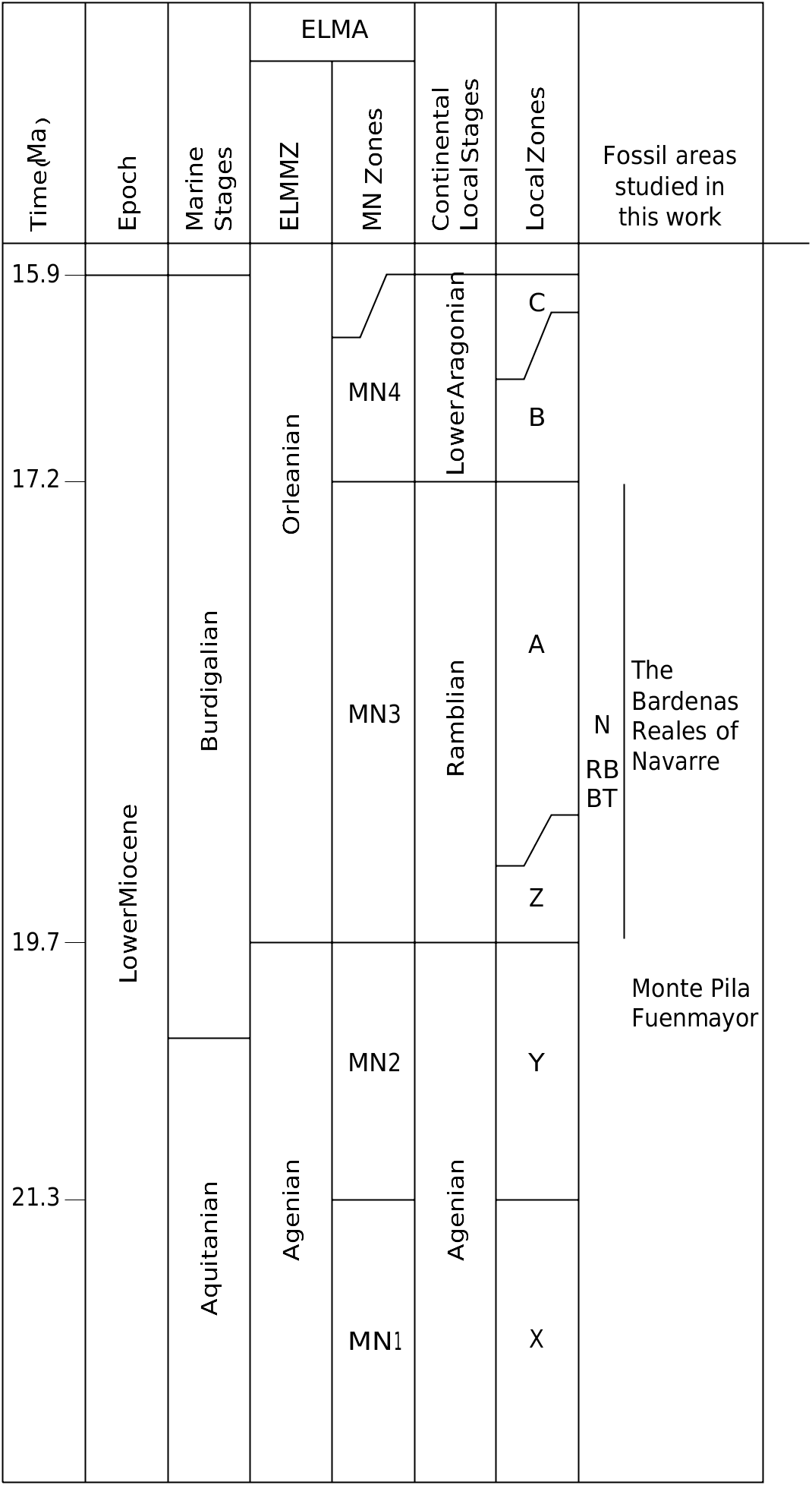
LOCALITY AND HORIZON. — Most specimens come from the Monte Pila sites, municipality of Lardero, Autonomous Community of La Rioja, Spain. Levels MP 1, MP 2 and MP 3 ( Figs 3-9 View FIG View FIG View FIG View FIG View FIG View FIG View FIG ). Other specimens came from the site of Fuenmayor, municipality of Fuenmayor, Autonomous Community of La Rioja, Spain ( Fig. 10 View FIG ). Western part of the Ebro Basin. Transitional facies between the Najera and Haro formations. Local zone Y, biozone MN 2, lower Miocene (see Figs 1 View FIG , 2 View FIG and the section Geographic and geological setting).
DESCRIPTION
The maximum length of the shells of the larger specimens of the freshwater turtles identified at both Monte Pila and Fuenmayor is close to 20 cm ( Figs 3-10 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG ). However, elements corresponding to individuals of very varied sizes are identified, some of them with a length of less than half that of the largest specimens. Some of the most complete shells, as well as the articulated elements of other specimens recognized as adult individuals due to their size, show fusion of many of their sutures, so that the margins of each plate cannot be recognized in them ( Figs 3 View FIG ; 5 View FIG ). The thickness of the plates is greater in the larger individuals. Although the width/length ratio of the shell is variable, this form shows a relatively wide carapace, being recognized as slightly longer than wide in some cases ( Fig.3G View FIG ). The carapaces are low.
The abundance of specimens allows assessment of the variability in morphology and dimensions of several elements. For example, the relatively wide nuchal plates are almost as long as wide in some individuals, but slightly wider than long in others ( Fig. 4 View FIG A-L). All of them show a thickening in the visceral region. The neural series anteriorly contacts the nuchal plate, and posteriorly the suprapygals. The neurals are wide in relation to their length ( Fig. 4 View FIG M-O; 10C, D). The morphology of these plates is variable. However, most of them are hexagonal, with the shortest margins being postero-laterally located. Although the anterior suprapygal is subquadrangular, the posterior is hexagonal, noticeably wider than long ( Fig. 4 View FIG Q-W). The pygal is also subquadrangular ( Fig. 4 View FIG X-Z).
This taxon has a single cervical scute ( Figs 3A View FIG ; 4 View FIG A-L; 9F). Its lateral margins are subrounded. Its width/length ratio shows a wide range of variability, being slightly wider than long in some specimens, but almost twice as long as wide in others. The overlap of the cervical on the visceral surface of the nuchal is relatively long. The vertebral scutes are hexagonal, almost as wide as they are long ( Fig. 3 View FIG A-G). The first and last ones are slightly wider than the second to fourth vertebrals, which are all similar in width. However, the vertebral series is narrow, the first scute being narrower than the nuchal in many cases, not overlapping the postero-lateral margins of this plate. The fifth vertebral overlaps the anterior region of the pygal plate ( Fig. 4 View FIG X-Z).
The contact between the hyoplastra and the hypoplastra, and that between the hypoplastra and the peripheral series, corresponds to a ligamentous junction, so that a hinge is developed between these plates, allowing mobility of the posterior plastral region ( Figs 3F View FIG ; 5 View FIG A-F; 7B-M; 8; 9D-E; 10A, B). The lateral margins of both plastral lobes are subrounded. The anterior lobe is very wide. Its anterior margin varies from subrounded to almost straight. Although the posterior lobe is clearly expanded in some specimens, it is much narrower in others. Thus, the development of the anal notch is variable in width and length, as is the shape of its margins ( Figs 3F View FIG ; 7 View FIG N-U; 8; 10A, B). The epiplastral lip is concave ( Figs 5 View FIG ; 6 View FIG A-V; 9A, B; 10A, B, G-L). Both the length and the thickness of this lip are observed to be highly variable. The dorsal thickening of this lip shows a pair of anterior processes, which exceed the margin of the anterior plastral lobe, at the level of the contacts between the gulars and the humerals.
The gular scutes overlap the anterior region of the entoplastron in some specimens, but not in others ( Fig. 6 View FIG W-AC). In the same way, the humero-pectoral sulci may be located next to the posterior margin of the entoplastron in some specimens, but overlap this plate in others, ranging from a very short overlap of the pectorals on the entoplastron in some individu- als, to a relatively long overlap, greater than a quarter of the length of this plate, in others. The abdominal scutes overlap the posterior region of the hyoplastra ( Figs 5 View FIG A-F; 7B-I; 9D-E; 10A). The anal scutes are restricted to the xiphiplastra, their anterior margin being well away from the suture between those plates and the hypoplastra ( Figs 7 View FIG N-U; 8; 9C; 10A, N).





| MP |
Mohonk Preserve, Inc. |
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.