
Diasporus ventrimaculatus, Chaves, Gerardo, García-Rodríguez, Adrián, Mora, Alejandra & Leal, Alejandro, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.187415 |
|
DOI |
https://doi.org/10.5281/zenodo.6215175 |
|
persistent identifier |
https://treatment.plazi.org/id/66487E5A-840D-FFF7-FF78-FF55FE74FA1D |
|
treatment provided by |
Plazi |
|
scientific name |
Diasporus ventrimaculatus |
| status |
sp. nov. |
Diasporus ventrimaculatus View in CoL sp. nov.
Holotype. Male, UCR 20493; collected on July 10, 2008 by Gerardo Chaves, Adrián García, Guillermo Artavia and Grupo de Guías Locales de Biolley y Altamira in the Valle del Silencio at the east edge of the Cordillera de Talamanca 20 km west from the Costa Rica-Panama border (9.1116 N, - 82.96172 W, 2550 m elevation).
Paratopotypes. Male: UCR 20494, UCR 20500, UCR 20503, UCR 20504, UCR 20505 UCR 20506; Female: UCR 20491 and UCR 20492. All material of these specimens were gathered by the same collectors and in the same place as the holotype.
Diagnosis. A member of the genus Diasporus based on the characteristic “dink” call, oval palmar tubercle, longitudinal gular folds in the edges of the deflated vocal sac and the presence of spadate disk covers on some fingers and toes. Members of this species have (1) skin on dorsum and venter areolate, but some individual are smooth; ventral disk absent; (2) tympanic membrane absent but tympanic annulus evident beneath skin, tympanum length is 50% of eye; (3) snout short, mucronate in dorsal view (truncate in females), truncate in profile; (4) upper eyelid lacking tubercles (but some individuals have few tubercles), as wide as interorbital distance (IOD); cranial crest absent; (5) vomerine odontophores absent (only present in the black male, mentioned below); (6) vocal slits large, external vocal sac present, nuptial pads absent; (7) first finger shorter than second, the last commonly with disk round, other fingers with disk spadate; (8) fingers broad, bearing narrow lateral fringes; subarticular tubercles round and flattened; palmar tubercle oval, round and flat, shorter than thenar tubercle; supernumerary tubercles not obvious (9) ulnar tubercle and fold absent; (10) heel and tarsus lacking tubercles and folds; (11) outer metatarsal tubercle flat and round; supernumerary tubercles not obvious (12) toes with lateral fringes; webbing absent; disk spadate (in juveniles slightly expanded); toe III reach the penultimate subarticular tubercle of toe IV; (13) light gray interorbital bar and black parietal area; dorsal head varies from cream to black; upper lip cream with small dark areas formed by small black dots; dorsum smooth red to pink in males, black with numerous small black dots in females; venter varies from red to pink in males and some of them with small white and black spots on ground color, otherwise females with large white blotches in a black ground; groin and underside area of limbs color vary according to the venter coloration in both sexes; posterior surfaces of thighs vary from red to black; dorsal limbs with diffuse transversal bars former by small dark dots, and in general these small black dots cover many areas of the body include bones; (14) SVL, in males 20.2 – 23.5 mm, in females 23.8 – 24.7 mm.
Holotype description. Adult male; body moderately robust; head about as wide as body, 116.7% wider than longer; head width 39.1% SVL; head length 33.5% SVL; snout round to truncate in dorsal view and truncate in profile; canthus slightly concave, softly sharp in profile; loreal region nearly flat; lips rounded; nostrils barely protuberant, directed laterally; width of upper eyelid less 81.6% of interorbital distance; eye large, its diameter much greater than its distance from nostril (123.8%); tympanum round with annulus visible beneath the skin, separated from eye by distance less than the tympanum diameter; supratympanic fold weak, running straight posteriorly, from point above tympanum, obscuring posteriodorsal edge of tympanum. Tongue small, transversally oval and it´s free for about one third of its length; choanae small, oval, directed laterally and cover by the maxillae in ventral view; vocal slits present, large, longitudinal, lateral to base of tongue.
Forelimb slender; ulnar tubercles and fold absent; palmar tubercle low, round and about same size as thenar tubercle; subarticular tubercles large and rounded; supernumerary tubercles absent; fingers slender, with small lateral fringes; relative lengths of fingers I<II<IV<III; tips of fingers expanded, rounded, and spadate in most of them; nuptial pads absent. Hind limb moderately slender; tibia length 50.2% SVL; foot length 40.8% SVL; heel and tarsus lacking tubercles and folds; metatarsal tubercle round and large; subarticular tubercles large and rounded; supernumerary tubercles absent; toes slender, with lateral and small fringes; relative lengths of toes I<II<III<V<IV; tips of toes expanded, even round, spadate, with small circumferential grooves. Skin on dorsum smooth; upper eyelids lacking tubercles; skin on venter areolate; discoidal fold not evident; skin ventral and ventrolateral to cloacae granular.
Coloration of holotype on preservative. Dorsum of head cream with diffuse darker brown spot to the anterior area; interorbital bar light gray and parietals are black, supratympanic folds outlined above and below by dark chocolate brown dots; dorsal surface of forelimbs slightly wither brown, with a small dark dots in the line of the wrist; dorsal surface of hind limbs similar to forelimbs with diffuse brown crossbars on shanks, thighs less colored but many small black dots; upper lip with diffuse brown bars; flanks lighter brown, changing ventrally and posteriorly to more red, with small white spots; groin red with small white spots and numerous small black dots; gular sac red with few and small black dots; ventral surfaces mottled with white spots on reddish ground color; iris pale brown.
Coloration of holotype in life ( Fig. 2 View FIGURE 2 BDE). Dorsum orange, with a light gray interorbital bar black parietals that turns into darker diffuse color that extent near to the middle of the body. The anterior part of the head is slightly more yellow than the rest of the body, with a thin black line that starts in the snout, crosses the eye and ends at the tympanum. Dorsal surface of the limbs are orange with black spots in the forearms and some of the digits; black spots in the hindlimbs above a less colored surface. Flanks orange; groin red with small white blotches. Ventral surfaces of body and limbs are red with big and irregular white spots, vocal sac uniform red. A black ring delimited orange iris.
Measurements of holotype (mm). SVL 23.3; tibia length, 11.7; head length, 7.8; head width, 9.1; interorbital distance 3.8; width of eyelid 3.1; diameter of eye 2.6; diameter of tympanum 1.3; fourth toe length 9.5.
Measurements of paratopotypes (mm). Measurements from six males and two females with mean and range in parenthesis. SVL males 21.8 ± 1.2 (20.2 – 23.5), females 23.9 ± 0.8 (23.2 – 24.7); tibia length, males 11.0 ± 0.7 (10.0 – 11.9), females 12.2 ± 0.0; head length, males 7.3 ± 0.3 (7.0 – 7.8), females 8.3 ± 0.1 (8.2 – 8.3); head width, males 8.4 ± 0.3 (7.9 – 8.7), females 9.5 ± 0.7 (9.0 – 10.0); interorbital distance, males 3.7 ± 0.2 (3.3 – 3.9), females 3.9 ± 0.3 (3.7 – 4.2); width of eyelid, males 3.2 ± 0.2 (2.9– 3.8), females 3.9 ± 0.1 (3.8 – 3.9); diameter of eye, males 2.7 ± 0.2 (2.3 – 3.1), females 3.2 ± 0.1 (3.1 – 3.2); diameter of tympanum, males 1.3 ± 0.2 (1.0 – 1.8), females 1.5 ± 0.1 (1.4 – 1.5); fourth toe length, males 8.9 ± 0.6 (7.8 –9.7), females 10.90 ± 0.42 (10.6 –11.2).
Color polymorphism and sexual dimorphism. Males ( Fig. 2 View FIGURE 2 B, 3C): dorsal head varies from cream to gray with a variable number and distribution of small black dots, a few individuals had red, brown or white blotches, all but one of the males present diffuse black and white bars in the upper lips; dorsum varies from red to brown with many small black dots that in some individuals form small dark blotches; groin red with small white blotches and small dark dots, but in some individuals are beige with or without white blotches; dorsal surface of thigh red with many brown blotches and small dark dots produce diffuse bars in most of the individuals; throat varies from orange to red with variable amount of small black dots; venter and underside thigh fluctuate from beige to red; all the individuals have small white blotches and have great variation in the number and distribution of the small black dots; iris red, but may be brass-colored in some individuals.
Black male ( Fig 3 View FIGURE 3 D): Dorsum black, snout and flanks are darkish gray. Venter and underside area of the limbs are similar to the females. This is the only male with female coloration.
Non-reproductive males ( Fig 3 View FIGURE 3 B): Head, snout, dorsum and tibia uniform reddish gray; interorbital bar light gray; throat similar to reproductive males; venter, groin and underside area of the thigh from beige to red with irregular large black and white blotches similar to females. Iris is brass colored.
Females ( Fig 2 View FIGURE 2 A, 3A): head, snout, dorsum and tibia uniform darkish gray or with white and black blotches; venter, groin and underside area of the thigh are black with irregularly sized and irregularly distributed white blotches that are bordered with many small black dots; iris is red or brass-colored.
Juvenile ( Fig 2 View FIGURE 2 C): SVL 18.3 ± 3.4 mm (n = 4). Head, snout, posterior area of dorsum and tibia uniform beige or with irregular yellow, red, brown and cream spots; interorbital bar yellow and parietals brown; throat, venter, groin and underside area of the thigh transparent with many small black dots and some beige and black blotches. Iris is brassy.
Hatchling: SVL 9.2 ± 2.0 mm (n = 2). Tip of the snout, interorbital bar and dorsum black separate with large white blotches. Flanks are black and white. Venter and underside area of the legs are whitish gray.
Molecular genetics. 364 nucleotides from the RAG-1 gene could be compared between individuals UCR 20491 (adult female), UCR 20496 (adult male) and UCR 20497 (juvenile male). Four SNPs could be identified. Sequences UCR 20491 and UCR 20496 were 99.725% identical (only one difference), sequences UCR 20491 and UCR 20497 were 99.176% identical (three differences), whereas sequences UCR 20496 and UCR 20497 were 98.901% identical (four differences). All codons, from all individuals, codify for the same aminoacids (no missense mutation have been found): all SNPs found are non-coding. On the contrary, alignment between the new species´sequence and the Diasporus hylaeformis RAG-1 sequence showed a transversion c.159 C>A ( EU186752 View Materials ) that means an aminoacid difference Thr53Asn (ABY57667) between the two species. Phylogenetic tree based on the RAG-1 sequences of these species and the most related ones found in the genomic on line database (BLAST), indicates that the new species and D. hylaeformis have a very close evolutionary history, and it is closer to the Caribbean genus Eleutherodactylus than to other mainland genus of direct development frogs such as Craugastor and Pristimantis .
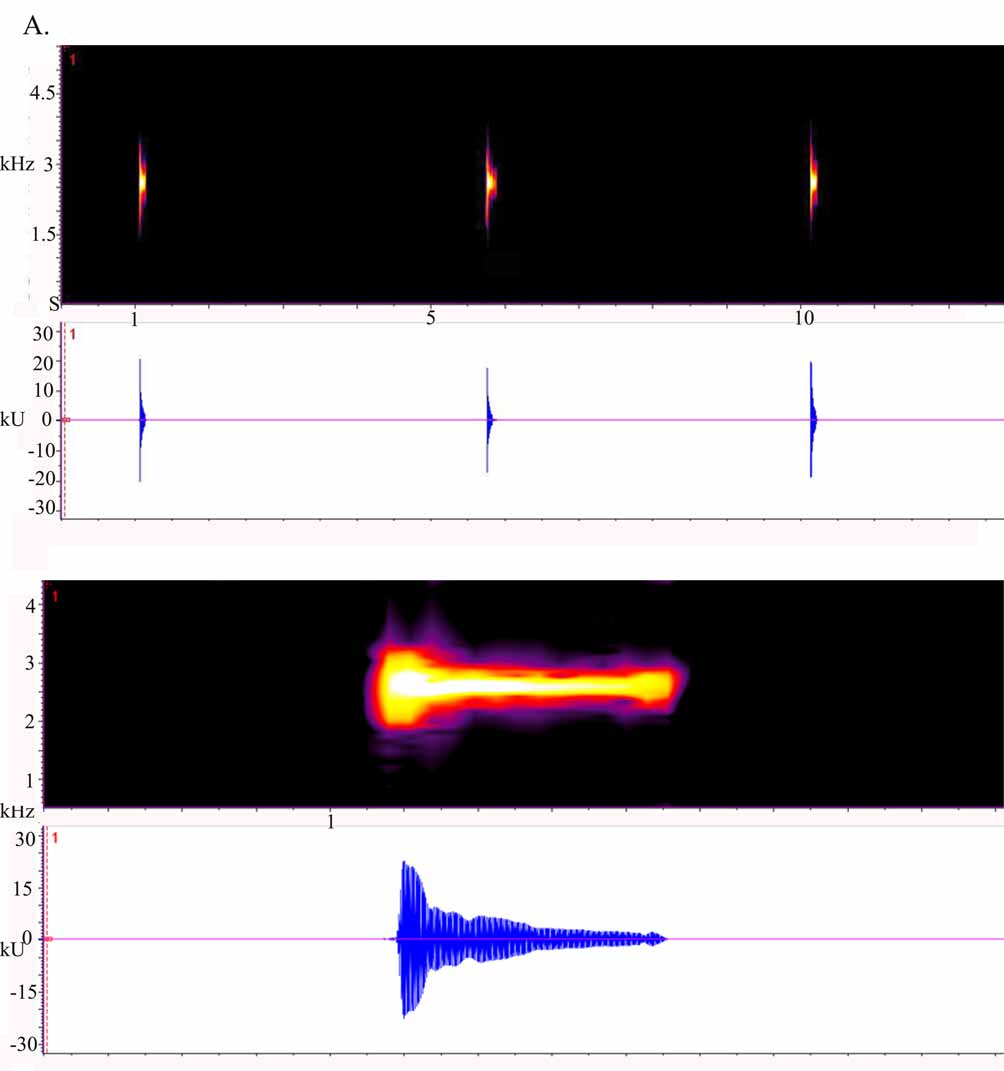
Vocalization. The call structure is given in Fig. 4 View FIGURE 4 . The mean temperature during recording was 14 ºC at 00:45 hours. Vocalizations produced by the holotype and other 5 males recorded and analyzed, are “dinks” similar to those of the other Costa Rican species of Diasporus ; a sound reminiscent of a note from a xylophone. The call consists of monophasic notes that are repeated slowly in a range between 2140 and 2995 Hz ( Fig 4 View FIGURE 4 A). The typical and dominant and fundamental frequencies ( Fig. 4 View FIGURE 4 B) are the same, about 2550 ± 55 Hz (2495–2605) and there is no evident frequency modulation. Call duration ( Fig 4 View FIGURE 4 B) is about 79.65 ms ± 8.90 ms and interpulse interval 5.15 s ± 1.55, with a call rate of approximately 11 pulses per minute.
Natural history. This species is found in many areas of the Valle del Silencio oak forest especially near clumps of terrestrial bromeliads. They can also be found in different layers of the forest, from canopy to less than half meter above the ground. We never observed them on the ground. Most males call inside the vegetation on the upper surfaces of the leaves, branches and stalks from both arboreal and terrestrial bromeliads and trees, but some individuals were exposed on the leaves close to the trails. All of the females where found in the vegetation, at least at 50 cm height, in generally lower than the males.
We did observe several males, females and juveniles of various sizes emerging in the leaf litter trapped between the leaves of the giant terrestrial bromeliad, Greigia sylvicola , in the beginning of the night (between 6 to 7 o’clock).We estimate that we heard hundreds of calling males in one particular small area of forest, where it was common to find more than one male perched in the same plant. Males start to call in the evening (18:00 to 19:00) until late in the morning (5:00 to 6:00) and can occasionally be heard during the day but in less frequency and abundance. On sunny days fewer males call than during rainy days and most of them call close to the flooded areas.
Distribution. On the trail between Altamira and Valle del Silencio (1300 to 2600 m), we found this species to occur around 2500 m of altitude. and only in the Valle del Silencio. We never detected D. ventrimaculatus on the slopes of the valley but only in the flat area. We did not detect individuals along a 3 km trail that runs along the east side of the Rio Terbi at the base of Cerro Asidbeta, this trail it´s only 15 meters from the holotype place but in the opposite margin of the river. Furthermore, this species was not present in the Jardín (9.1298 N, - 82.9673 W, 2450 m), a larger open moss-backed area with numerous treelike ferns (fog) located on the northeastern part of the valley. Near Cerro Bekón (9.1594 N, -83. 0 478 W, 2600 m), 11 km to west of the Valle del Silencio and at the same altitude, the senior author never saw any individuals of the new species during a survey in December 2007, in spite of surveying more than one hundred terrestrial bromeliads.
Numerous surveys were conducted in this region in the late 20th century by noted herpetologists including William Duellman, Charles W. Myers, Douglas Robinson, Federico Bolaños, Karen Lips, David Wake and others ( Duellman & Myers 1980; Lips 1998; Federico Bolaños personal communication). These surveys included Cerro Pando (8,9250 N, - 82.7269 W, 2400 m), Mellizas (8.8895 N, - 82.7775 W, 1400 m), Sitio Las Tablas (8.9361 N, - 82.7528 W, 1880 m), Alturas de Cotón (8.9392 N, - 82.8390 W, 1400 m) and Cerro Echandi (9.0340 N, - 82.8246 W, 3100 m) and in spite of these past surveys D. ventrimaculatus was not reported or collected at these localities. In the 1980’s in a survey carried out by the courageous Costa Rican botanist Luis Diego Gómez to Cerro Bine (9.0950 N, - 82.8929 W, 3242 m), at the headwater of the Río Terbi, he not registered this species there, although he did collect an important new species of salamander there (Federico Bolaños personal communication).
Based on these historic surveys we infer that this species probably does not occur outside of the southeastern region of the Valle del Silencio, specifically in the flat and flooded area of around 2.5 km ² in the west side of the Río Terbi ( Fig. 1 View FIGURE 1 ), enclosed by the Hoffmann, Sin Nombre, Turubo and Adsibeta peaks and the moss-backed area of the Jardín.
Conservation status. We calculated an EVS score of 17 for Diasporus ventrimaculatus that indicates a maximum conservation priority. In spite of the abundance of Diasporus ventrimaculatus and the good conservation condition of its habitat, we believe that this high EVS value and its reduced geographical extension of less than 3 km ² put this species in a critical endanger category under the IUCN criterion B2aciv. We hypothesize that the number of adult reproductive individuals would fluctuate very much in a small population and there are several examples of drastic amphibian population declines in other species with a reduced or small range of distribution in Costa Rica (see below for details in the discussion).
Etymology. The specific name ventrimaculatus is a composed term derived from the Latin words ventris meaning venter and maculatus meaning spotted. The name is in relation to the larger dark and white blotches in the venter found in most of the individuals and mainly in the females, this coloration is not found in any other species of the genus.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.