
Micruroides euryxanthus, AND OTHER ELAPIDS
|
publication ID |
https://doi.org/10.1206/3972.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5452930 |
|
persistent identifier |
https://treatment.plazi.org/id/6746F70E-5B61-FFCF-F636-FCFEFD0AF9BF |
|
treatment provided by |
Carolina |
|
scientific name |
Micruroides euryxanthus |
| status |
|
COMPARISONS BETWEEN M. EURYXANTHUS AND OTHER ELAPIDS
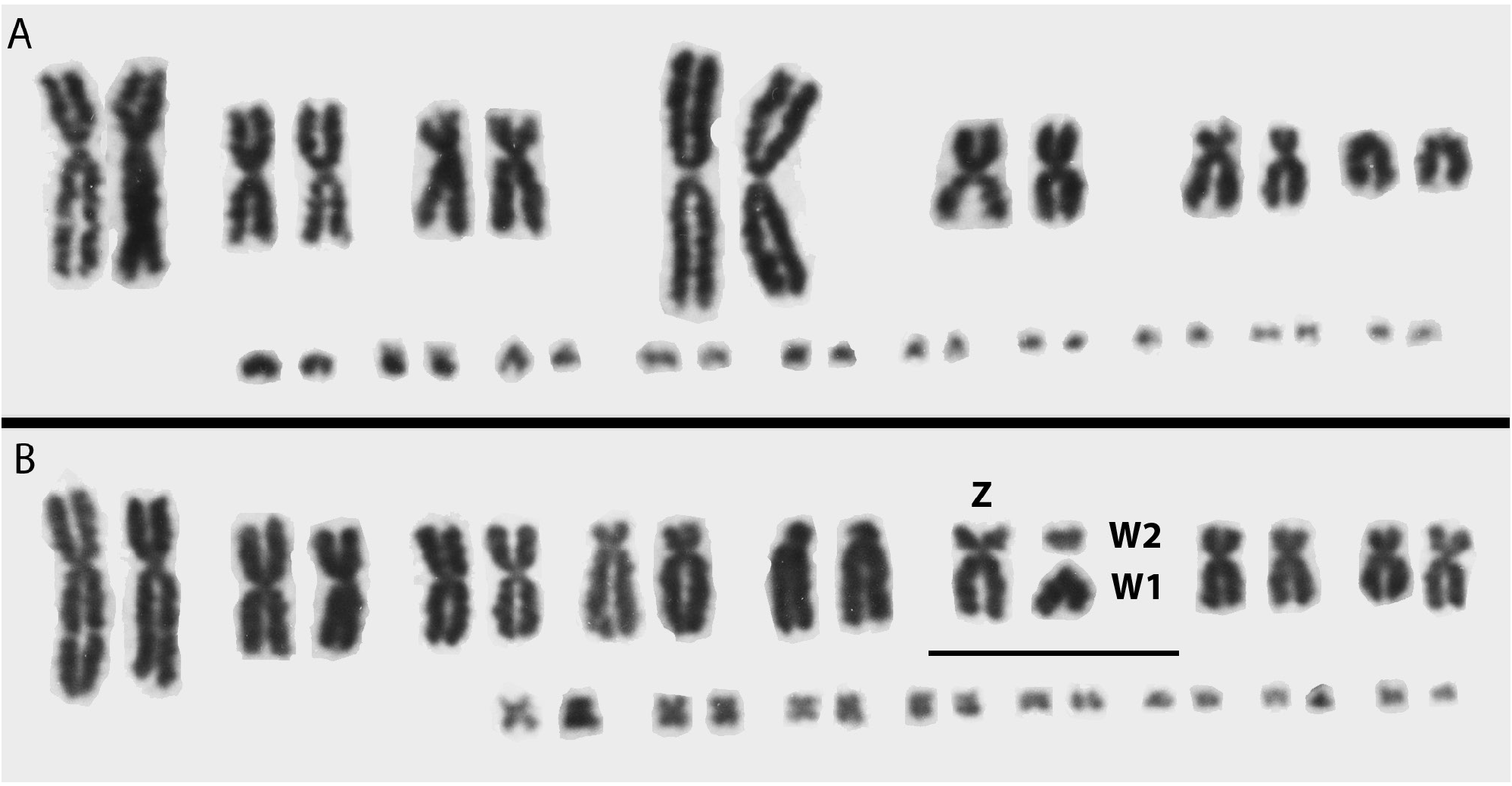
None of the karyotypes reported for elapids from the New World represents the ancestral karyotype in all details (2 n = 36, with 16 macrochromosomes and 20 microchromosomes). However, the karyotype of M. euryxanthus (2 n = 34, with 14 macrochromosomes and 20 microchromosomes) is close to this condition, with only one fewer pair of macrochromosomes. In addition, the first five pairs of macrochromosomes appear to be very similar to those in the common, ancient snake karyotype as found in many members of the Colubridae (reviewed by Cole and Hardy, 2019). In particular, pair 1, the largest metacentric chromosome in M. euryxanthus , appears to be the same as pair 1 in the ancestral karyotype, and this chromosome has been found in only two other species of coralsnakes. It occurs in the South American Micrurus surinamensis (see Gutiérrez et al., 1988) and the Central American Micrurus mipartitus (see Gutiérrez and Bolaños, 1979), but none of the other coralsnakes of the Western Hemisphere that have been karyotyped. In fact, the karyotype of M. mipartitus is basically identical to that of Micruroides euryxanthus . This is consistent with the phylogenetic conclusion ( Slowinski, 1995; Castoe et al., 2007) that Micruroides euryxanthus represents the earliest clade of New World coralsnakes that is the sister taxon to all the others, and suggests that the presence of the large metacentric macrochromosome is retained from the ancestral condition. The presence of apparently the same macrochromosome in M. surinamensis and M. mipartitus may also be from retention of the ancestral condition, and its absence from other New World coralsnakes may reflect a historic transformation by means of centric fission to being represented in the other species as in M. tener ( fig. 1B View FIGURE 1 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.