
Korkonterpeton kalnense, Werneburg & Štamberg & Steyer, 2020
|
publication ID |
https://doi.org/ 10.37520/fi.2020.019 |
|
persistent identifier |
https://treatment.plazi.org/id/6B5B87F8-CE35-FFE5-8220-FEA81F84FA18 |
|
treatment provided by |
Felipe |
|
scientific name |
Korkonterpeton kalnense |
| status |
sp. nov. |
Korkonterpeton kalnense sp. nov.
Text-figs 3–11 View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig
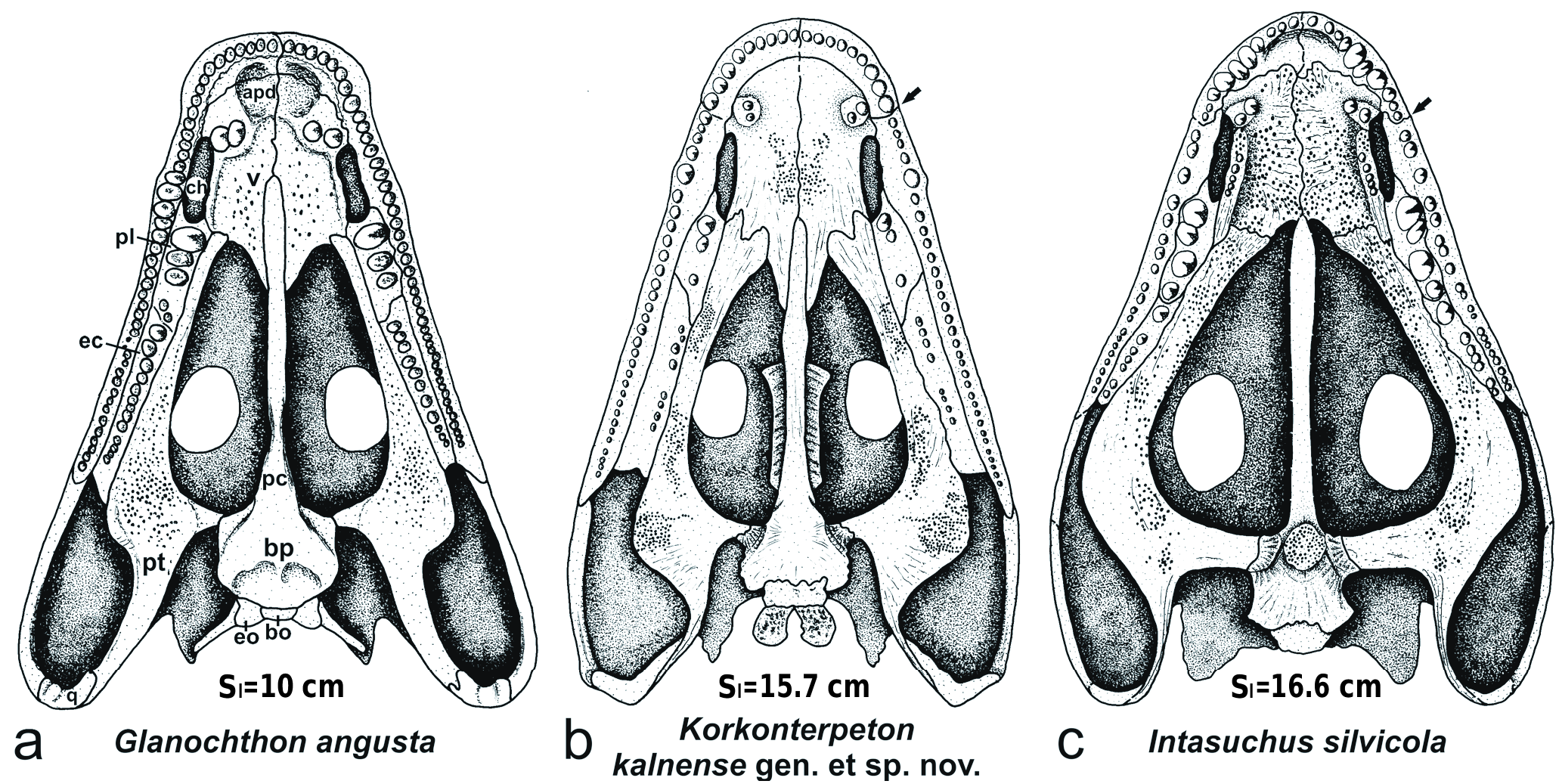
H o l o t y p e. MEBHK – P 82447, a subcomplete skeleton corresponding to an adult individual (see below), and preserving a skull (length 15.7 cm) in dorsal and ventral views, and well preserved but disarticulated postcranial elements.
T y p e h o r i z o n. Pinkish laminated claystone of the upper Kalná Horizon, upper part of Prosečné Formation (Asselian to Artinskian, lower Permian).
T y p e l o c a l i t y. Outcrop ‘Track in ravine on the edge of the forest’ (coordinates 50° 33′ 58.599″ N; 15° 43′ 18.873″ E) near the village of Arnultovice , 3 km north of the town Hostinné, Krkonoše Piedmont Basin, Bohemia GoogleMaps , the Czech Republic.
E t y m o l o g y. After the finding layer of the upper Kalná Horizon, named after the main fossil locality Horní Kalná, near Arnultovice.
D i a g n o s i s. Stereospondylomorph with the following unique character combination:
(1) Tabular extremely elongated, as long as the supratemporal.
(2) Posteromedian skull margin extremely concave, with very distant posterior tabular corner (Thl/Sl = 0.14).
(3) Postparietal very short, half the length of that in related stereospondylomorphs.
(4) Posterior margin of the quadratojugal and dorsomedial quadrate concave, with a steplike shelf region.
(5) Cultriform process of the parasphenoid slightly swollen in its mid-region, its anterior third embedded in the posteromedian region of the vomer.
(6) Sphenethmoid and epipterygoid ossified early.
(7) Well-ossified, ventrally large hook-shaped quadrate, with an elongated anteromedial part.
Additional characters in contrast to those in Cheliderpeton vranyi , but shared with a few other stereospondylomorph temnospondyls:
Additional characters in contrast to those in a few stereospondylomorph species, but shared with others: (14) Posterior quadratojugal enclosing the posterior part of the squamosal (may be similar in C. vranyi and Intasuchus ).
(15) Dorsal part of the quadrate forming a narrow anteromedial directed bony-strip with a posterior process-like boss as the most posterior part of the skull (shared with archegosaurids and eryopids).
(16) Posterior process of the vomer very elongated almost up to the level of the posterior region of the palatine (similar in Glanochthon ).
(17) Ectopterygoid posteriorly wide (shared with Archegosaurus ).
(18) Pterygoid with very short basipterygoid (shared with Glanochthon and Archegosaurus ).
(19) Anterior palatine ramus of the pterygoid medially curved (shared with Archegosaurus and Intasuchus ).
(20) Transversal process of the pterygoid laterally swollen (similar in Glanochthon and Sclerocephalus ).
(21) Basal plate of the parasphenoid short (similar in Glanochthon and Sclerocephalus ).
(22) Anterior tip of the parasphenoid cultriform process reaching the level of the anterior portion of the pterygoid (shared with Archegosaurus and Intasuchus ).
(23) Basipterygoid process of the basal plate anterolaterallydirectedandsuturedwiththebasipterygoid process of the pterygoid (similar in Intasuchus ).
(24) basal plate ventrally smooth (shared with Glanochthon and Sclerocephalus ).
D e s c r i p t i o n. Preservation. The skeleton of Korkonterpeton kalnense gen. et sp. nov. is well preserved and has been fully prepared: its skull is large (compared to the size of the postcranial elements) and visible in dorsal and palatal views. The hemimandibles are also visible from both sides, labially and lingually. The postcranial skeleton is disarticulated but with well-preserved elements including the vertebral column (with the atlas) and ribs.
Growth stage. The skull has a midline length of 15.7 cm, and shows clear dermal ornamentation consisting of large sub-circular polygons in growth zones, turning into well-marked ridges and grooves, towards the periphery of the bones. The endocranium is also well ossified (see below). Together with the fact that the ribs present uncinate processes and the vertebrae transverse processes (see also description below), this indicates that MEBHK – P 82447 corresponds to an adult individual ( Steyer 2000b).
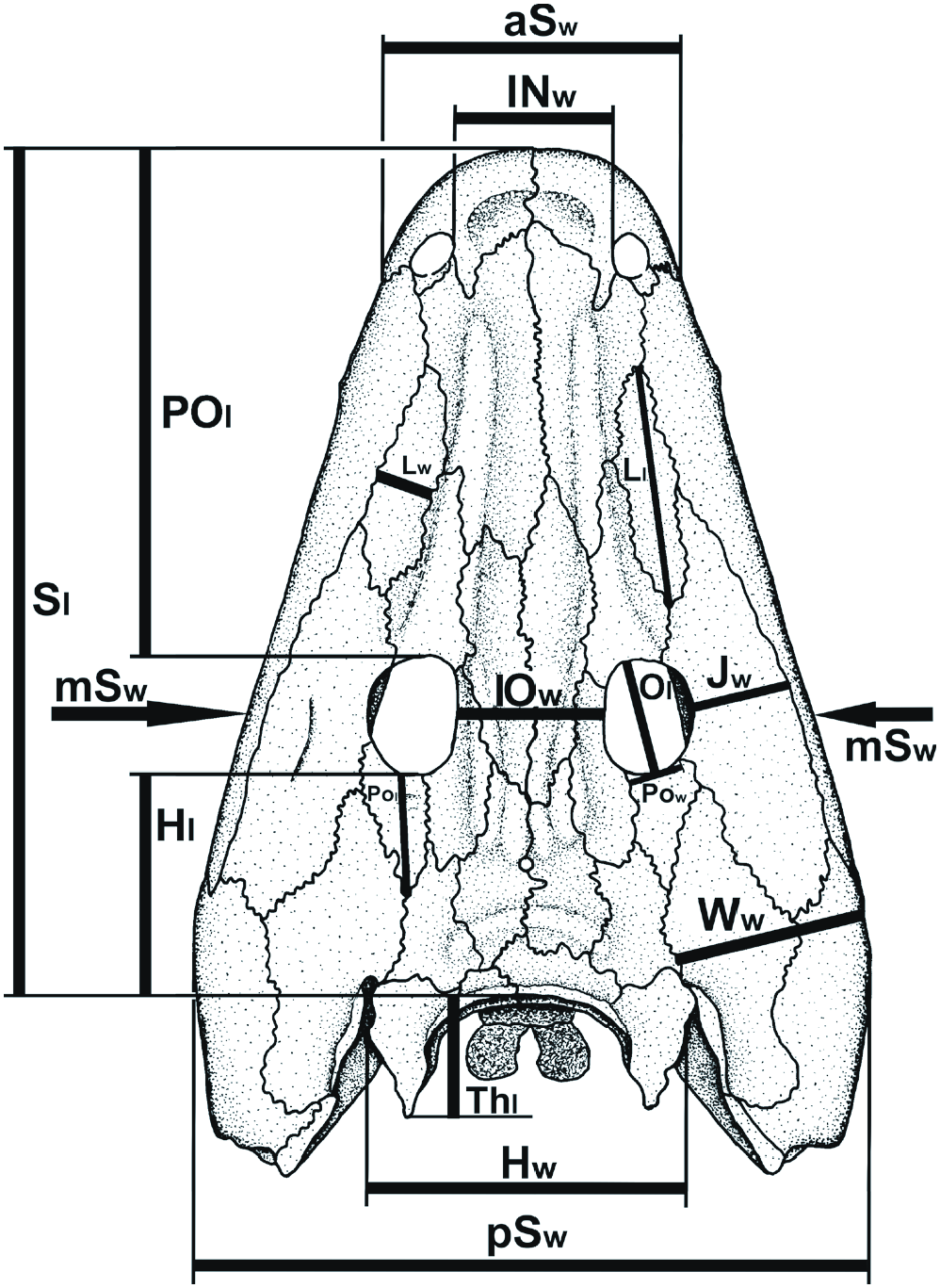
Skull roof ( Text-figs 3–5 View Text-fig View Text-fig View Text-fig , 14 View Text-fig , Tab. 1). The skull is subtriangular in outline, with a very elongated preorbital region (2.45 times the skull table length; Tab. 1: POl/Hl = 2.45; POl/ Sl = 0.60) and a short postorbital skull table (Hl/Sl = 0.26), in contrast to C. vranyi . The tip of the snout is relatively wide (aSw/Sl = 0.35), box-like and blunt, as in Glanochthon ( Schoch and Witzmann 2009b) and Sclerocephalus ( Boy 1988, Schoch and Witzmann 2009a). The intranarial distance (INw/Sl = 0.19) is longer than the intraorbital (IOw/ Sl = 0.17). The premaxilla is relatively short medially but it presents posteriorly a very elongated alary process that goes beyond the posterior margin of the external nostril. The septomaxilla is visible dorsally, but its dorsal contribution is very small. The lateral margin of the skull roof, bordered by the maxilla, is very straight except at the level of the anterior tip of the lacrimal where the maxilla presents a small lateral expansion to accommodate a large tooth. This is sometimes the case in Glanochthon ( Schoch and Witzmann 2009b: fig. 2). In Korkonterpeton kalnense gen. et sp. nov., the maxilla is elongated and posterodorsally reaches the quadratojugal. Its suture with the nasal is relatively long, as in Glanochthon angusta ( Schoch and Witzmann 2009b) . The lacrimal is relatively narrow and does not make contact with the nostril nor the orbit. The nasal is anteriorly widened and anteriorly extends to the anterior margin of the naris (in contrast to Intasuchus ). The intraorbital region (IOw/ Sl = 0.17) and the frontal are narrow. The orbits are subcircular and relatively small compared to the skull size (Ol/ Sl = 0.14). The prefrontal is anteriorly narrow and very elongated: it contacts the lacrimal in its anterior half. The jugal is relatively narrow, in contrast to Intasuchus .
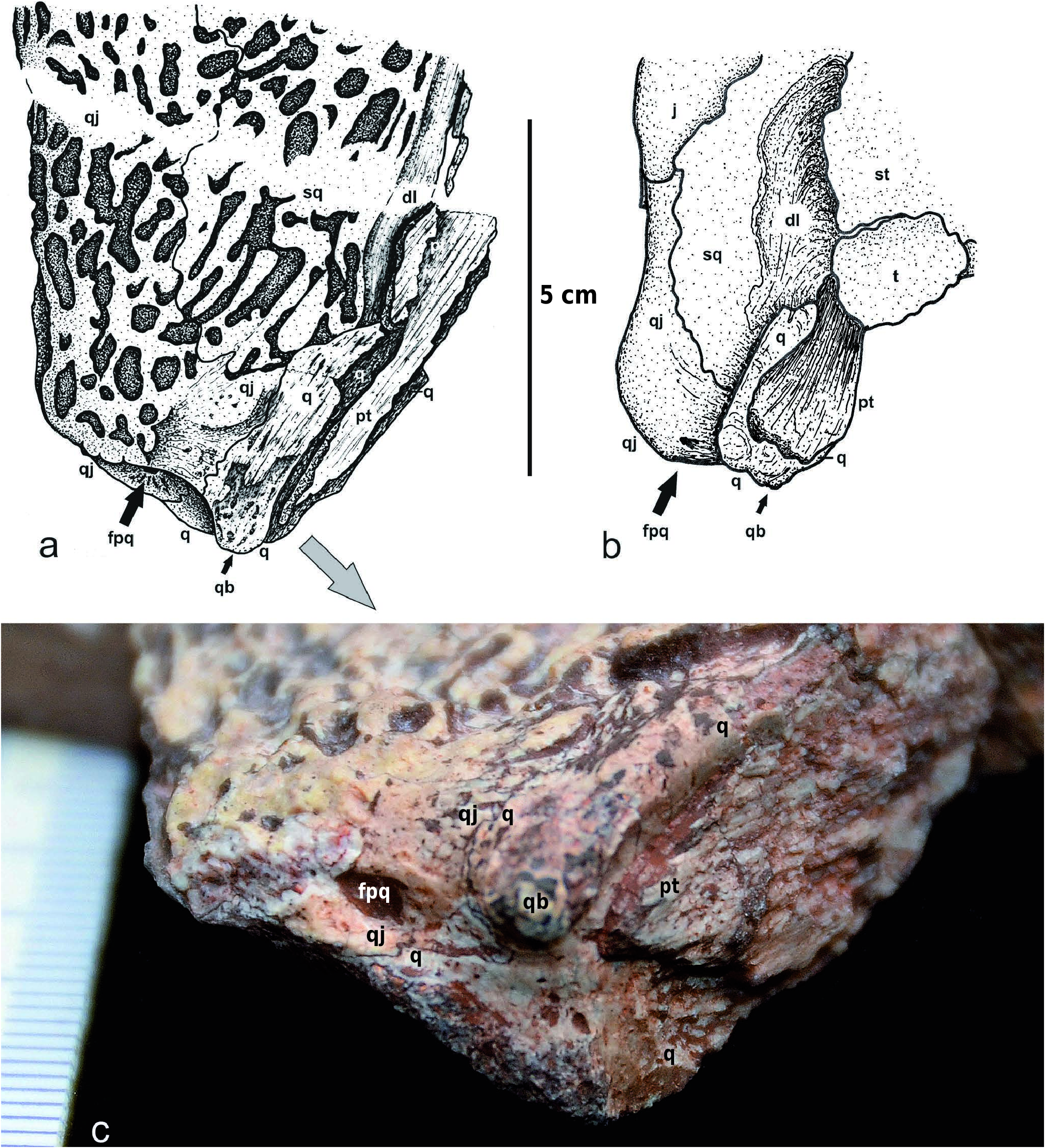
The maximum width of the skull roof is at the level of the anterior portion of the quadratojugal (pSw/Sl = 0.79), as in Intasuchus but in contrast to Glanochthon and Cheliderpeton vranyi ( Werneburg and Steyer 2002) . The quadrate is visible dorsally: its small dorsal contribution lies posterolateraly to the otic notch, and it contacts the quadratojugal laterally, the descendent lamina of the squamosal anteriorly, and the vertical pterygoid wall medially ( Text-fig. 5a View Text-fig ). The posterodorsal portion of the quadrate forms the quadrate boss. This boss is also visible in the archegosauroids Platyoposaurus stuckenbergi (PIN 3968/1; RW pers. obs.), Melosaurus uralensis (MB.Am.334a; RW pers. obs.), Kashmirosaurus ornatus ( Werneburg and Schneider 1996: fig. 2F) and in the eryopid Clamorosaurus borealis (PIN 3950/1; RW pers. obs. and Gubin 1983: 114). In Korkonterpeton kalnense gen. et sp. nov., the quadrate boss, together with the posterodorsal portion of the quadratojugal, borders a very deep and concave condyle region of the skull roof. This region is so deep that the paraquadrate foramen is partly visible dorsally ( Text-fig. 5a View Text-fig ), as in the aquatic eryopid Glaukerpeton ( Text-fig. 5b View Text-fig and Werneburg and Berman 2012). This condyle region is also significantly posterior to the occiput.
The postorbital is narrow and elongated, extending more posteriorly than the postfrontal. The postfrontal anteriorly contacts the prefrontal. Its posterior extremity reaches the level of the pineal foramen. The parietal and supratemporal are relatively narrow and elongated, but the supratemporal is slightly shorter than that of Glanochthon or C. vranyi . In Korkonterpeton kalnense gen. et sp. nov., this supratemporal does not contribute to the otic notch, as in Glanochthon angusta but not Intasuchus silvicola .
Thepostparietalisveryshort.Thetabularisveryelongated (as long as the supratemporal). The posterior margin of the skull table is extremely concave (Thl/Sl = 0.14). These characters were unknown within stereospondylomorphs so far.
The dorsal skull roof shows an interesting strutting pattern composed of distinct ridges ( Text-fig. 3 View Text-fig ): the prefrontal ridge (prr, Text-fig. 3b View Text-fig ), the most developed, is longitudinal. It starts narrowly at the level of the premaxillar alary process and runs along the nasal and prefrontal where it widens posteriorly. This ridge is known in many temnospondyls (e.g., Sawin 1941,
pm – premaxilla, po – postorbital, pp – postparietal, prf – prefrontal, prr – prefrontal ridge, psr – parietal-supratemporal ridge, pt – pterygoid, ptf – postfrontal, q – quadrate, qj – quadratojugal, sm – septomaxilla, sq – squamosal, st – supratemporal, t – tabular. Sl – midline skull length.
Boy 1993, Werneburg 2007, Werneburg and Berman 2012). The intraorbital ridge (ior, Text-fig. 3b View Text-fig ) is also longitudinal. It starts at the level of the pre- and postfrontal suture, runs along the suture between the frontal, the pre-and postfrontal, and ends posteriorly on the anterior part of the parietal, as in C. vranyi ( Text-fig. 14e View Text-fig ), but in contrast to Glanochthon latirostris ( Schoch and Witzmann 2009b: fig. 4B). It is not clear if it is a continuation of the prefrontal ridge. The parietal-supratemporal ridge (psr, Text-fig. 3b View Text-fig ) is transversal: it runs anteriorly along the parietal-postparietal suture and laterally reaches the supratemporal, as in Intasuchus (Textfig. 14c) and eryopids ( Sawin 1941, Boy 1990, Werneburg 2007, Werneburg et al. 2010). In Korkonterpeton kalnense gen. et sp. nov., the premaxillar ridge is also transverse and runs anteriorly along the premaxilla-maxilla suture. Only the postparietal ridge is very narrow and typically runs along the posterior skull table margin.
The lateral line sulci are not well developed in Korkonterpeton kalnense gen. et sp. nov.: only the narrow infraorbital sulcus is relatively marked but partly visible only on the postorbital and suborbital regions (ifc.pt unique deeply marked and ifc.sb respectively, Text-fig. 3b View Text-fig ).
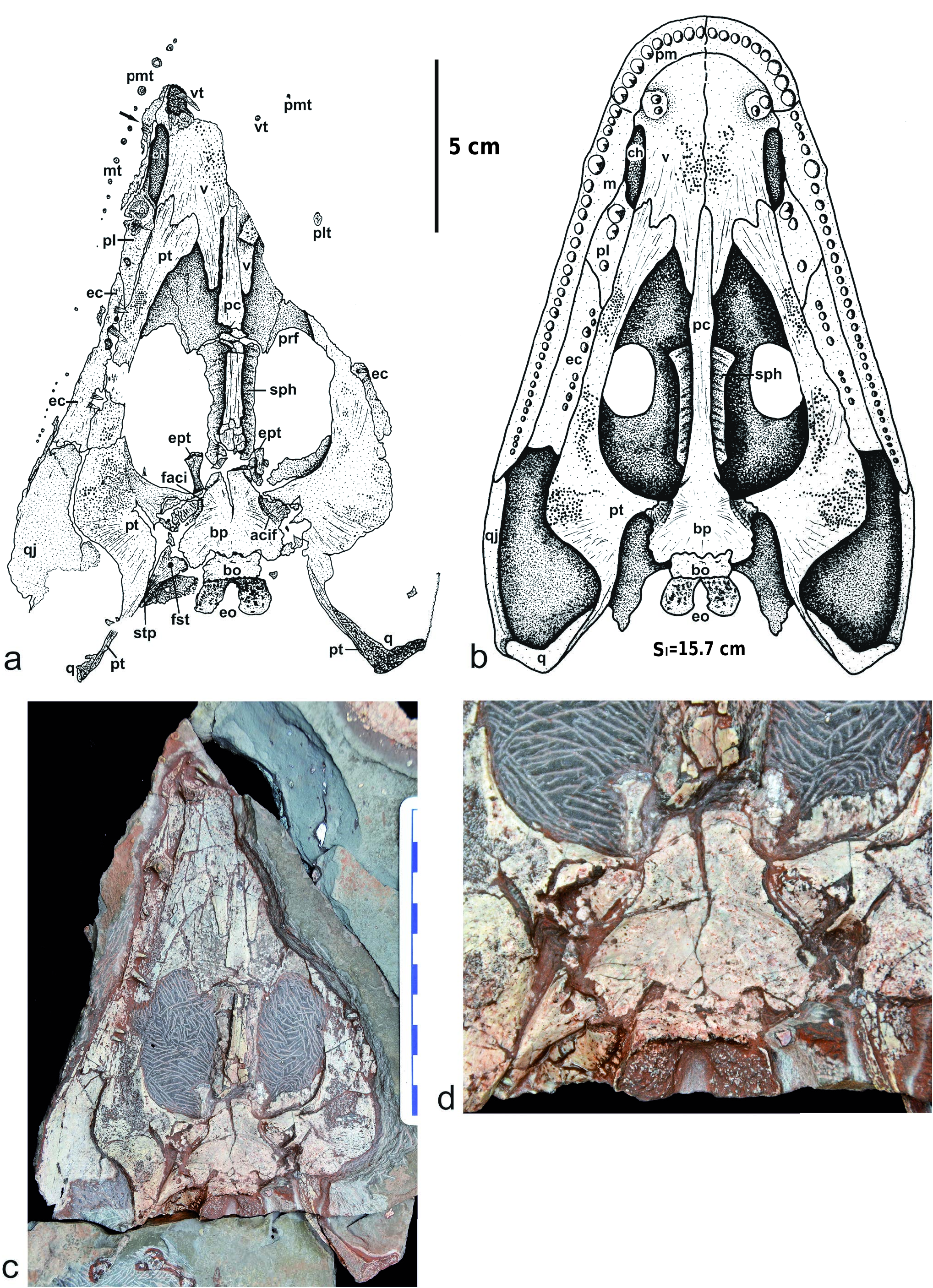
Palate ( Text-figs 6 View Text-fig , 15 View Text-fig ). The palate is well preserved and reveals interesting details concerning the dentition: the premaxilla bears about nine teeth, the three last ones being the largest – they are also larger than the anterior teeth of the maxilla. The fifth tooth of the maxilla, at mid-length of the choana, is a tusk that is supported by a small lateral expansion of the bone. Posteriorly to this tusk, are about 25 maxillary teeth decreasing progressively in size. Medially, at the level of the choana, the maxilla also widens, as in Intasuchus .
The vomer is a very large and developed bone, covering most of the palatal surface of the snout. The vomerian tusks are double and located in a circular small depression surrounded by a bony crest. Surprisingly, they are anterior to the choana, and even anterior to the premaxilla-maxilla suture, a unique character within stereospondylomorphs. The vomer and the pterygoid are covered by numerous small denticles arranged in clusters. The posteromedial process of the vomer is very elongated along the cultriform process of the parasphenoid: it reaches the level of the posterior tip of the palatine, as in Glanochthon but not in Intasuchus .
The choana is very narrow and slit-like, as in all stereospondylomorphs ( Text-fig. 15 View Text-fig ), but it is narrower than that of Sclerocephalus and shorter than that of Archegosaurus .
The ectopterygoid, twice the length of the palatine, bears 8–10 relatively small teeth, except for the third and fourth which have the same diameter as the vomerian tusks (Textfig. 6a–c). The posterior width of the bone is similar to the anterior width, as in Archegosaurus (Witzmann 2006: fig. 3b), but its posterior part is wider than in Glanochthon and Intasuchus ( Text-fig. 15a, c View Text-fig ).
The palatine is of similar length to the choana. It bears 3–5 teeth, the two anterior ones being the palatine tusks. These tusks are however smaller than those of Glanochthon or Intasuchus .
The pterygoid has a very short basipterygoid ramus in contrast to Intasuchus . Its quadrate ramus is ventrally very narrow and sutures posteriorly with the very elongate quadrate. The palatine ramus is curved medially and its anterior extremity is very wide (2/3 the width of the vomer) and bilobed: its anterolateral lobe is anterior to the posterior tip of the choana. This palatine ramus is similar in proportion to that of Intasuchus , but much wider than that of Glanochthon . The transversal process of the pterygoid is laterally swollen.
The parasphenoid has a relatively short basal plate in comparison with other stereospondylomorphs. The cultriform process is narrow but slightly widens at half its length, posteriorly to the vomer and anteriorly to the sphenethmoid. The anterior third of the cultriform process is embedded in the vomer (contra the anterior half in Sclerocephalus and Glanochthon ; Schoch and Witzmann 2009a, b). The anterior tip of the cultriform process reaches the same level as the posterior extremity of the choana (in Sclerocephalus and Glanochthon , it reaches half the length of the choana). In Korkonterpeton kalnense gen. et sp. nov., the basipterygoid process of the parasphenoid is located anterolaterally on the basal plate, as in Intasuchus but not in Glanochthon nor in Sclerocephalus . At the base of this basipterygoid process, a fine groove for the carotid artery is visible: it ends anteriorly by the carotid foramen. The basal plate of the parasphenoid is ventrally smooth as in Glanochthon , but not in Intasuchus .
Braincase ( Text-fig. 6 View Text-fig ). The sphenethmoid and epipterygoids are well ossified (the epipterygoids are missing at this ontogenetical stage in Glanochthon and Archegosaurus ; Witzmann 2006, Schoch and Witzmann 2009b). The sphenethmoid lies dorsally above the cultriform process: it has an elongate rhombic shape with an anteriorly wider region, as in Sclerocephalus haeuseri GOLDFUSS, 1847 (where this bone is still shorter; Schoch and Witzmann 2009a: fig. 6B) but not in the eryopids (where it is arrowhead-like; Sawin 1941, Werneburg 2007, Werneburg and Berman 2012).
The epipterygoid ( Text-fig. 6a, c–d View Text-fig ) has a broad base, a poorly ossified end and a slender shaft. The unpaired basibranchial is similar in shape but much larger than the epipterygoid (the basibranchial is recorded in Glanochthon , Sclerocephalus and Archegosaurus ; see Witzmann 2006, Schoch and Witzmann 2009a, b). The basioccipital is relatively short and sutures the basal plate of the parasphenoid at the two-third region of its posterior margin. The exoccipital, which forms the occipital condyle, is very elongate posteriorly, a unique character within stereospondylomorphs.
The quadrate is ventrally a large hook-shaped and wellossified bone. Its medial suture with the quadrate ramus of the pterygoid is very elongate and oblique. As described above, this bone dorsally presents a quadrate boss.
Visceral skeleton ( Text-fig. 6a, c, d View Text-fig ). The stapes is preserved in the right otic notch: it has a wide footplate (width 8 or 13 mm) and an apparently relatively short and slender shaft (recorded length 19 mm). The quadrate process is not visible here (contra Sclerocephalus and Archegosaurus ; Witzmann 2006, Schoch and Witzmann 2009a).
Mandible ( Text-fig. 7 View Text-fig ). The mandible is well preserved, with its left hemimandible complete: it is relatively long (length = 205 mm) and shallow (height = 37 mm) compared with others stereospondylomorphs. The surangular is clearly visible but its suture with the angular remains difficult to detect precisely. The dorsolabial portion of this surangular is straight and very slightly ornamented. The surangular process is very flat. The articular is a relatively small bone (length = 18 mm) within the complete hemimandible. Dorsolingually, it forms an elongated and deeply concaved joint socket ( Text-fig. 7c View Text-fig ). Just ventrally to this socket, the articular-prearticular suture runs anteroposteriorly straight in lingual view. The foramen for the chorda tympani lies ventrally to this suture, in the posterior portion of the prearticular: this foramen is very shallow but relatively elongate (length = 10 mm). Labially, the angular and the postsplenial are well ornamented. The mandibular sulcus is
tooth, pm – premaxilla, pmt – premaxillary teeth, prf – prefrontal, ps – parasphenoid, pt – pterygoid, q – quadrate, qj – quadratojugal, sph – sphenethmoid, stp – stapes, v – vomer, vt – vomerian tooth. Sl – midline skull length.
clearly visible in the posterior region of the hemimandible, in labial view it consists of a smooth straight groove at the level of the angular-surangular suture. Three other larger grooves are also visible on the labial surface of the surangular: they represent the accessory mandibular sulcus, as in Archegosaurus decheni (Witzmann 2006: figs 21, 22). The splenial is relatively short (length = 22 mm) in labial view. The dentary is the longest bone of the hemimandible (length = 173 mm). It is slightly ornamented labially. The left dentary bears 21 preserved teeth, whose general size decreases posteriorly along the lower jaw. However, the anterior third and fourth teeth are smaller than the posterior ones. The largest tooth of the most anterior dental exhibits an anterior blade. In conclusion, the mandibular anatomy of Korkonterpeton kalnense gen. et sp. is very similar to that of the others stereospondylomorphs (e.g., Boy 1993, Schoch and Witzmann 2009a, Witzmann 2006).
Postcranium ( Text-figs 8–11 View Text-fig View Text-fig View Text-fig View Text-fig ). The postcranial skeleton is only represented by its axial skeleton. The anterior vertebrae and ribs are well preserved. The atlas-axiscomplex is especially completely recorded ( Text-figs 8–9 View Text-fig View Text-fig , 10a, b View Text-fig ). The atlas consists of a transversely narrow intercentrum fused to the neural arch. The notochord opening dorsally divides the centrum into two lateral lobes with the exoccipital facets in anterior view, as in Platyoposaurus stuckenbergi ( Gubin 1991: fig. 27). The thin low neural spine of the atlas consists of two processes that do not meet dorsally. A rudimentary transverse process is exposed on the concave lateral flank of the neural arch, as in Sclerocephalus ( Schoch and Witzmann 2009a) . The postzygapophysis is substantial for articulation with the axis. The prezygapophysis of the atlas is very distinctively exposed with an oval shape in anterior view. The proatlas is also preserved: its small neural arch has a globular bellied shape ( Text-figs 8 View Text-fig , 9 View Text-fig ). It was clearly in articulation with the atlas, as in Eryops ( Moulton 1974) , Sclerocephalus ( Boy 1988, Schoch and Witzmann 2009a) and Archegosaurus ( Witzmann and Schoch 2006) .
The axis bears a very robust neural spine, which is much more massive in anterior or posterior view than that of the atlas. It has an enlarged dorsal top and strong transverse processes. The first anterior rib articulates with the axis: this rib is very short and shows no clear uncinate process. The second rib articulates with vertebrae III and is more elongated than the first rib. It has enlarged ends and a very small hook-like uncinate process ( Text-fig. 9 View Text-fig ). The third, fourth and following anterior ribs are larger and have particularly elongated uncinate processes. Usually 13 anterior thoracic ribs with large uncinate processes are known in Sclerocephalus ( Boy 1988) , but this number may be lower in Korkonterpeton kalnense gen. et sp. nov. The posterior ribs, shorter and narrower than the anterior ones, have no processes. The neural arch of vertebra III (Textfig. 11a) shows oval and sculptured elements on the lateral side of its neural spine. The top of this spine is greatly enlarged as a transversal process. This transversal process laterally presents the diapophysis and a pleurocentral facet in posterior view. The vertebrae V–VIII also have expanded transversal processes and very narrow neural spines. The lateral flanks of these spines are smooth. The dorsal top of the spines is extremely expanded and forms a large oval, rough surface with a rugose margin ( Text-fig. 11b View Text-fig ). The smaller and more posterior vertebrae are illustrated in lateral view in Text-fig. 10c–e View Text-fig . The centra are well ossified. The intercentra are large and form robust crescent shaped elements. Their dorsal surface is rugose, whereas their ventral and lateral surfaces are smooth and slightly concave. The parapophyses are fully ossified. The paired, narrow-oval pleurocentra have smooth and concave lateral flanks. The pattern of these centra is clearly rhachitomous and very similar to those of stereospondylomorphs and eryopids ( Moulton 1974, Boy 1988, 1990, Witzmann and Schoch 2006, Werneburg 2007, Schoch and Witzmann 2009a,).
The ventral scalation is poorly preserved. Only a few spindle-like ventral scales are recorded ( Text-fig. 9 View Text-fig , below the right quadratojugal).
ComparisonwithMemonomenosdyscriton S T E E N, 1 9 3 8. Another stereospondylomorph anterior skeleton is known from the Czech Krkonoše Piedmont Basin: Memonomenos dyscriton STEEN, 1938, from the early Asselian Rudník Horizon (Vrchlabí Formation) of Košťálov, 36 km west of Trutnov. Milner (1978), Schoch and Milner (2000) and Witzmann (2006) placed this species in the genus Archegosaurus , later Schoch and Witzmann (2009b) established the validity of the genus Memonomenos as well as its close relationships with Intasuchus . However, this species most likely belongs to the Archegosauridae , not to Intasuchidae according to our observations (see Tab. 1): its posterior and middle skull width, its posterior skull table, its jugal/cheek and nostrils are indeed narrow, as is the case in the Archegosauridae . However, Memonomenos dyscriton differs from Archegosaurus decheni in having a wider anterior skull, a smaller ratio between the pre- and postorbital length (POl/Hl = 1.91), a shorter lacrimal, more elongated nostrils, and a slightly wider jugal. Two characters of M. dyscriton may be unique: the similar intranarial and intraorbital distances (INw/Sl = IOw/Sl = 0.13), and the relatively narrow jugal (Jw/Sl = 0.06).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |